

Transcriptomic taxonomy and neurogenic trajectories of adult human, macaque, and pig hippocampal and entorhinal cells
- PMID: 34798047
- PMCID: PMC8813897
- DOI: 10.1016/j.neuron.2021.10.036
Transcriptomic taxonomy and neurogenic trajectories of adult human, macaque, and pig hippocampal and entorhinal cells
Abstract
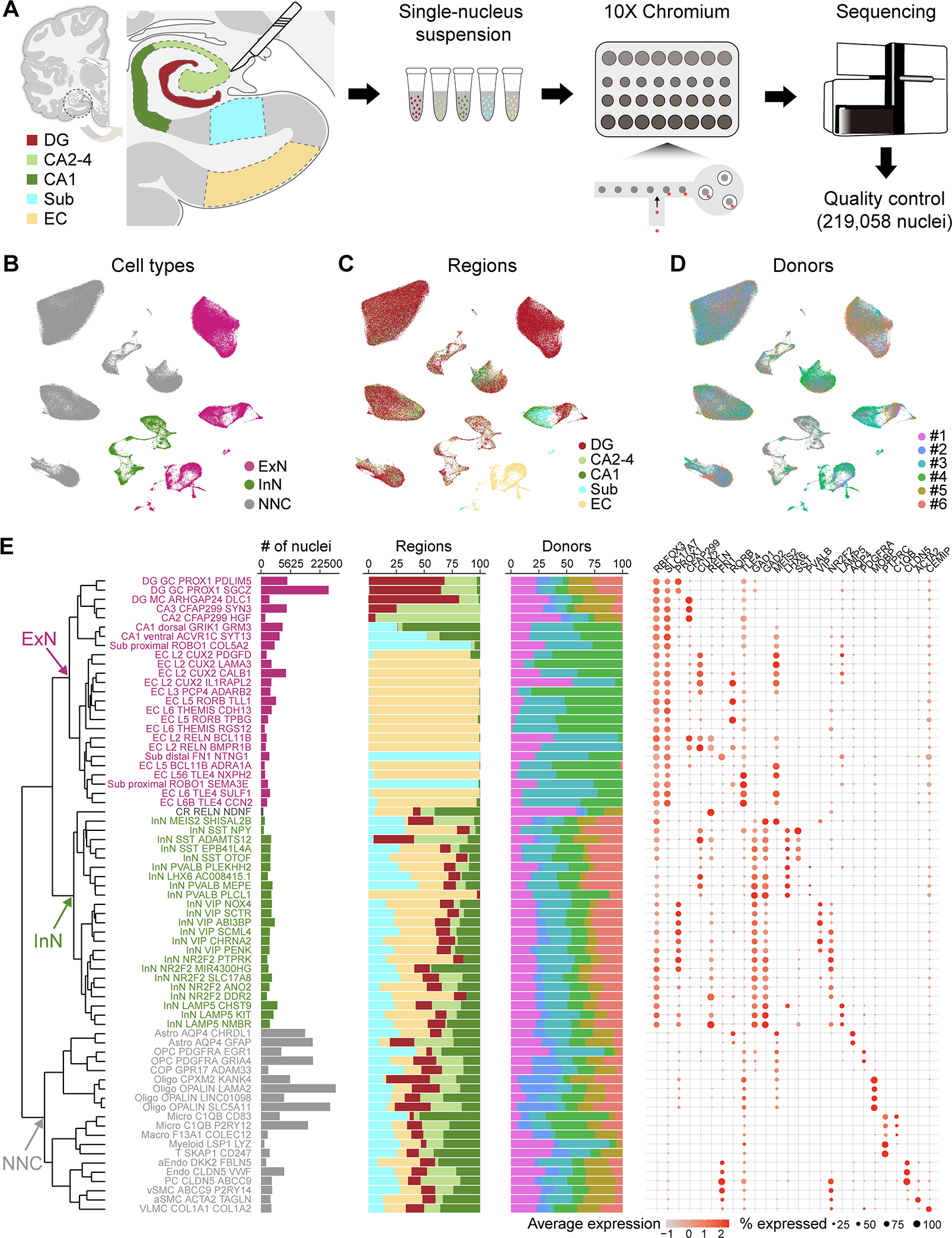
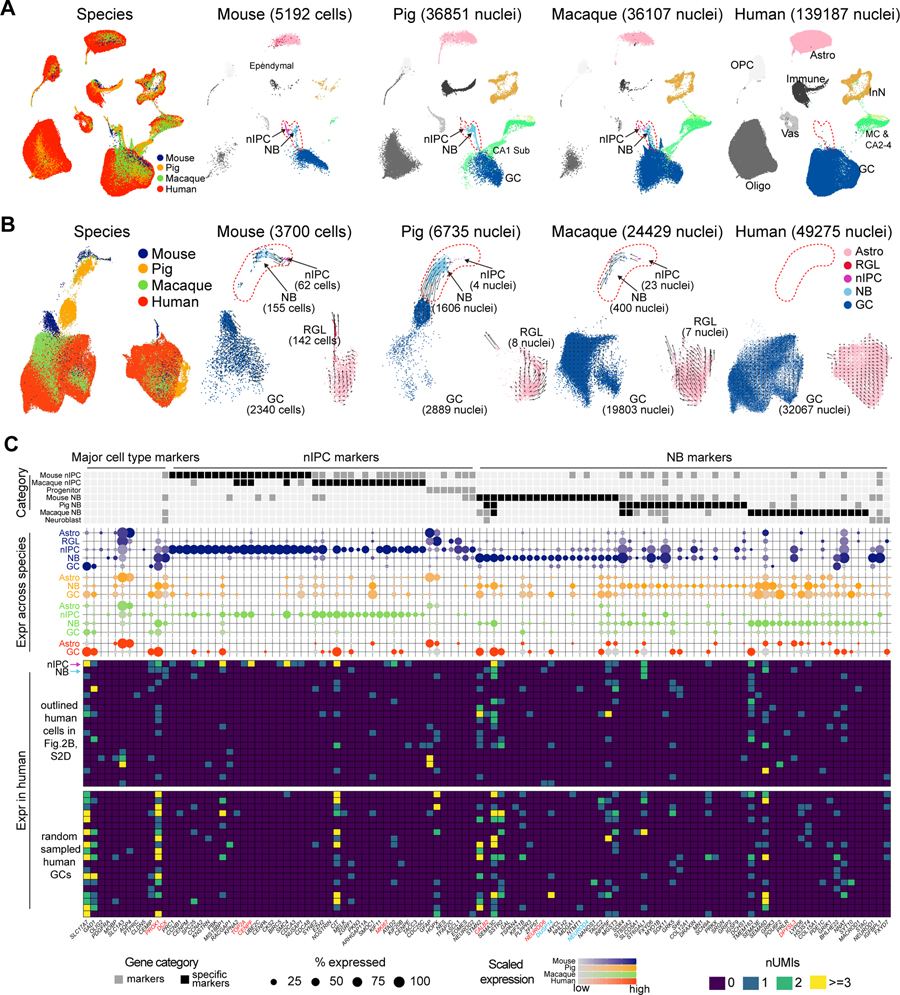
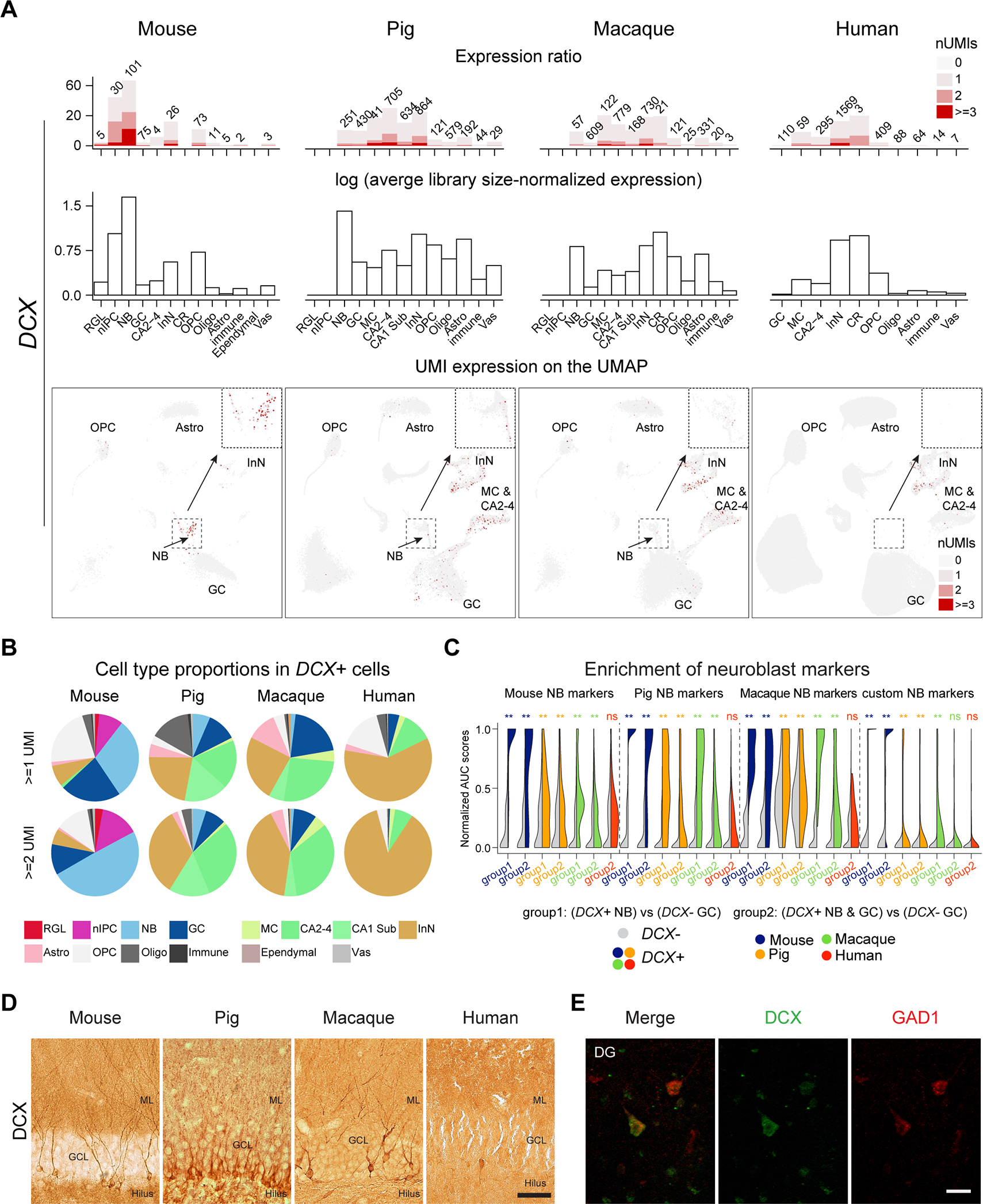
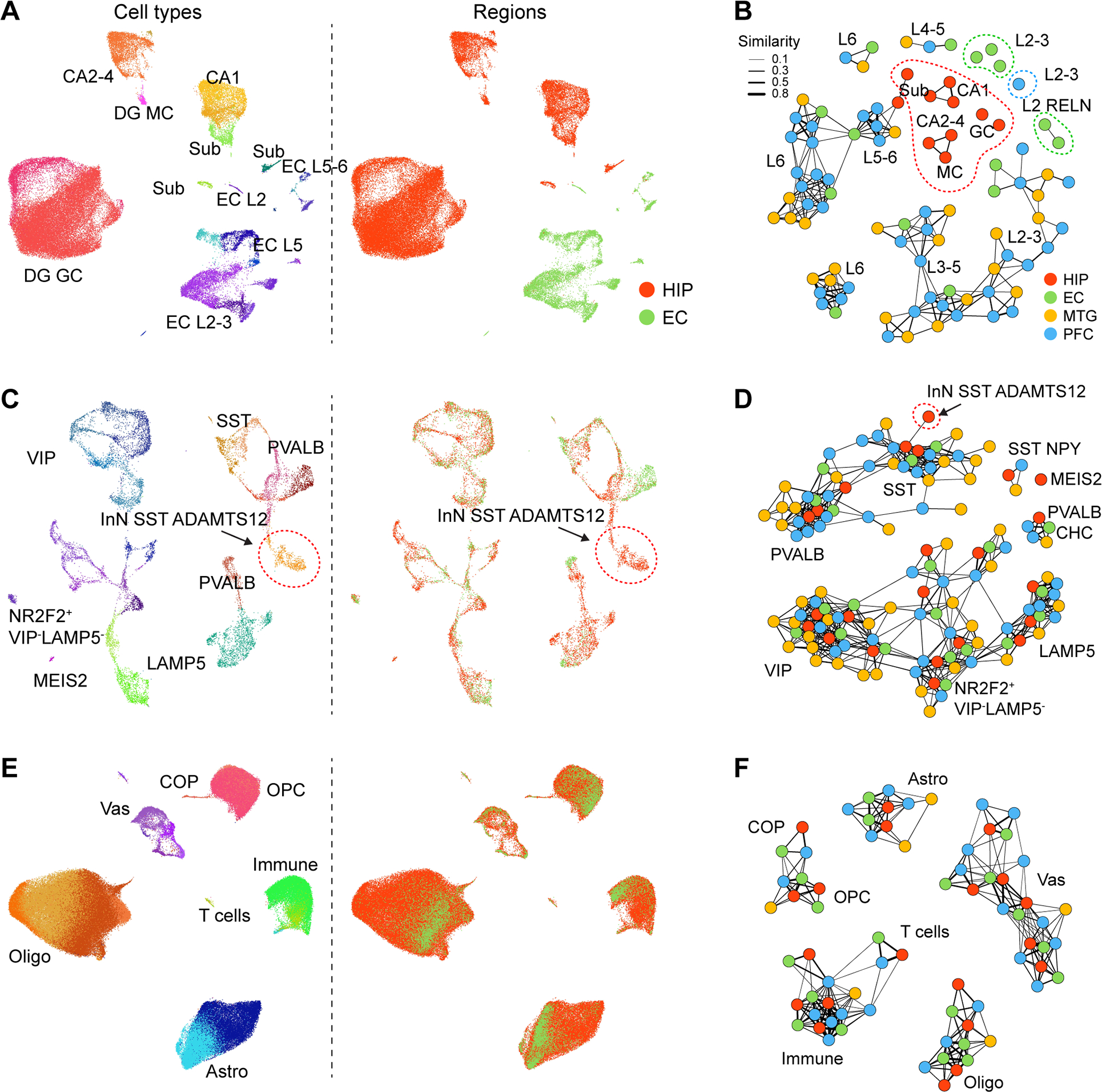
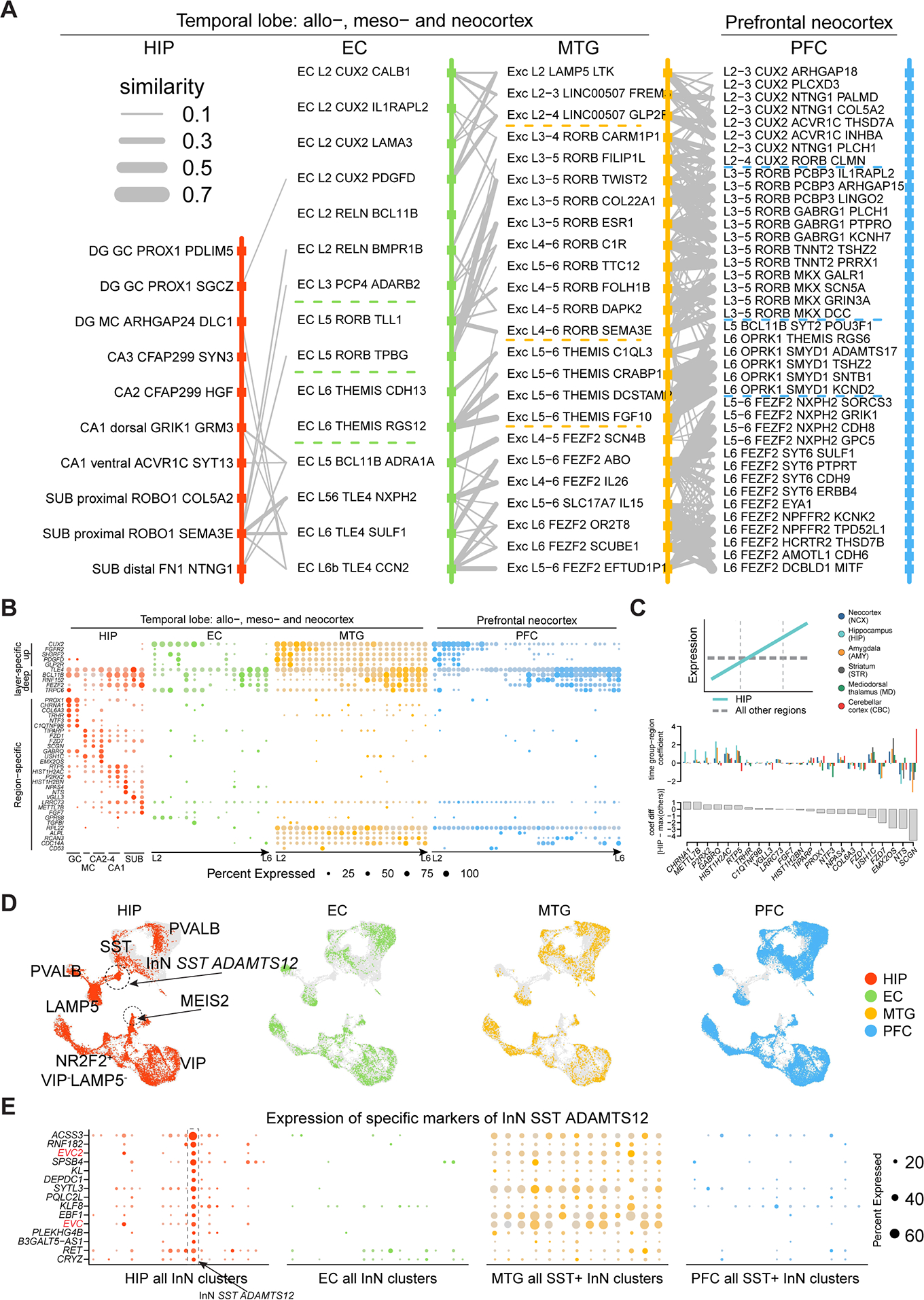
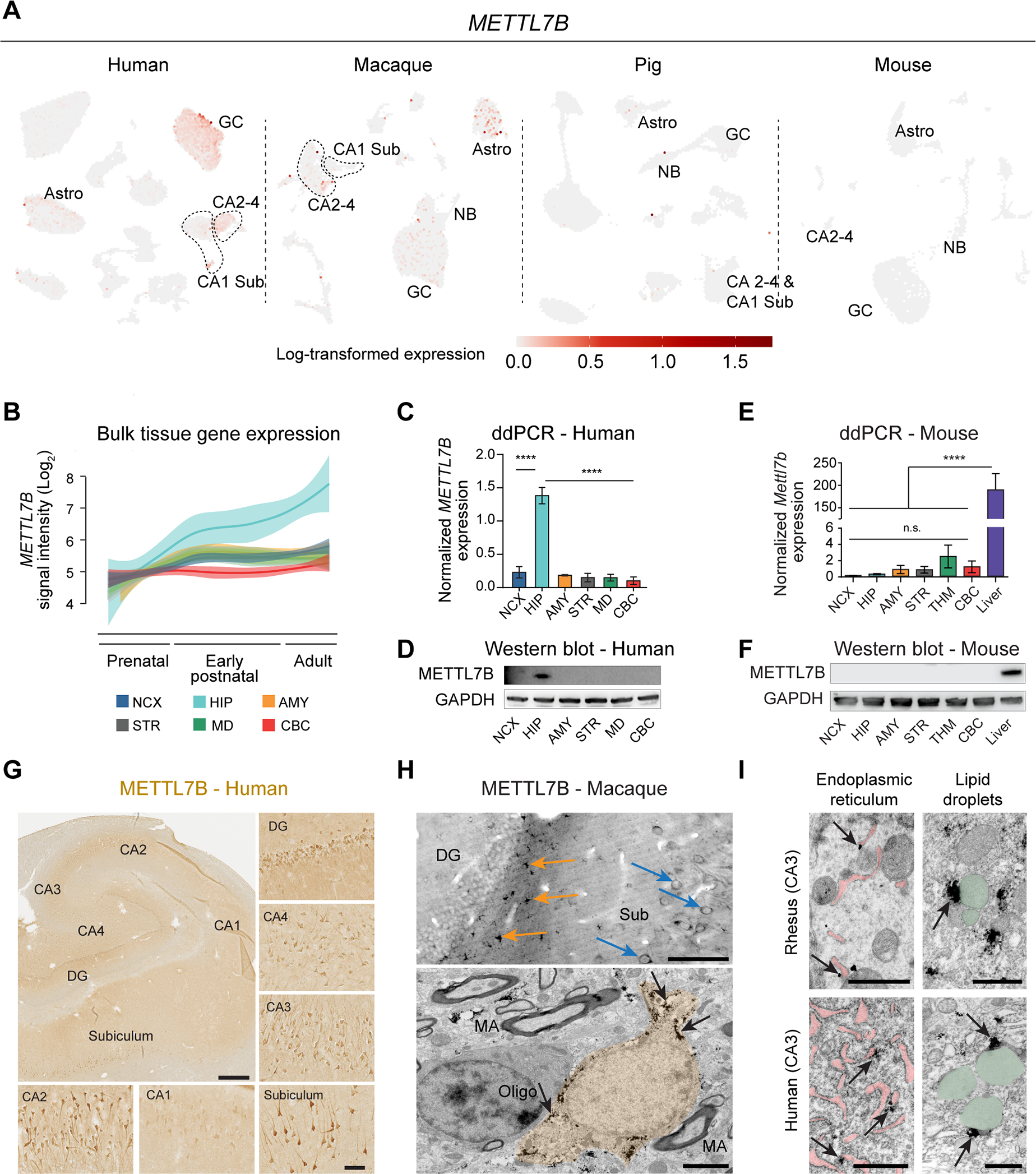
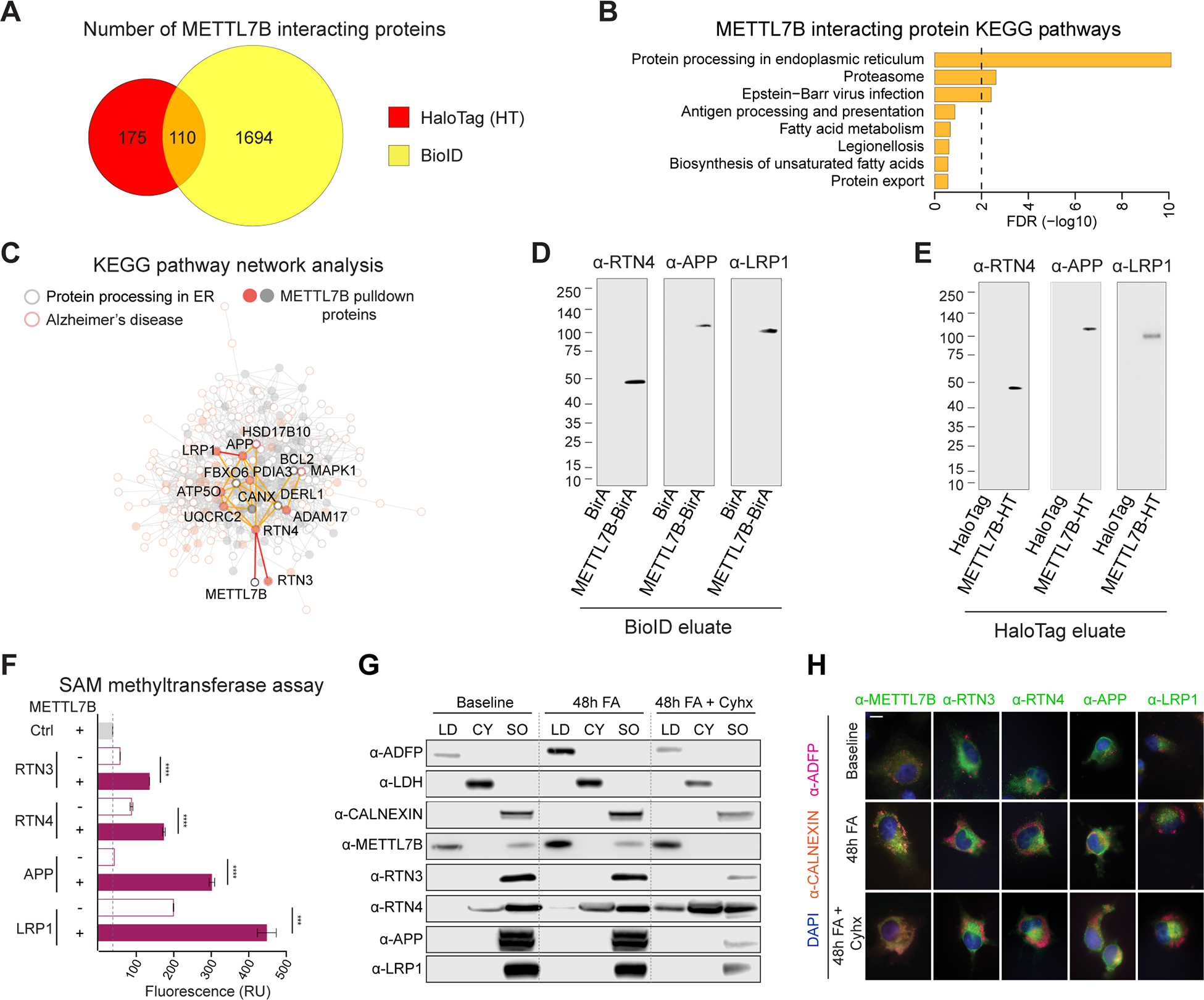
The hippocampal-entorhinal system supports cognitive functions, has lifelong neurogenic capabilities in many species, and is selectively vulnerable to Alzheimer's disease. To investigate neurogenic potential and cellular diversity, we profiled single-nucleus transcriptomes in five hippocampal-entorhinal subregions in humans, macaques, and pigs. Integrated cross-species analysis revealed robust transcriptomic and histologic signatures of neurogenesis in the adult mouse, pig, and macaque but not humans. Doublecortin (DCX), a widely accepted marker of newly generated granule cells, was detected in diverse human neurons, but it did not define immature neuron populations. To explore species differences in cellular diversity and implications for disease, we characterized subregion-specific, transcriptomically defined cell types and transitional changes from the three-layered archicortex to the six-layered neocortex. Notably, METTL7B defined subregion-specific excitatory neurons and astrocytes in primates, associated with endoplasmic reticulum and lipid droplet proteins, including Alzheimer's disease-related proteins. This resource reveals cell-type- and species-specific properties shaping hippocampal-entorhinal neurogenesis and function.
Keywords: Alzheimer’s disease; METTL7B; adult neurogenesis; doublecortin; entorhinal cortex; evolution; hippocampus; immature neurons; neuroblast; single-cell RNA-seq.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Single-nucleus sequencing finds no adult hippocampal neurogenesis in humans.Nat Neurosci. 2022 Jan;25(1):2. doi: 10.1038/s41593-021-00991-6. Nat Neurosci. 2022. PMID: 34992287 No abstract available.
-
Mounting evidence suggests human adult neurogenesis is unlikely.Neuron. 2022 Feb 2;110(3):353-355. doi: 10.1016/j.neuron.2022.01.004. Neuron. 2022. PMID: 35114104
References
-
- Abbott LC, and Nigussie F (2020). Adult neurogenesis in the mammalian dentate gyrus. Anat Histol Embryol 49, 3–16. - PubMed
-
- Andersen P (2007). The hippocampus book (Oxford; New York: Oxford University Press; ).
-
- Angata K, Suzuki M, and Fukuda M (2002). ST8Sia II and ST8Sia IV polysialyltransferases exhibit marked differences in utilizing various acceptors containing oligosialic acid and short polysialic acid. The basis for cooperative polysialylation by two enzymes. J Biol Chem 277, 36808–36817. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
