

The Smad4-MYO18A-PP1A complex regulates β-catenin phosphorylation and pemigatinib resistance by inhibiting PAK1 in cholangiocarcinoma
- PMID: 34799729
- PMCID: PMC8990017
- DOI: 10.1038/s41418-021-00897-7
The Smad4-MYO18A-PP1A complex regulates β-catenin phosphorylation and pemigatinib resistance by inhibiting PAK1 in cholangiocarcinoma
Abstract
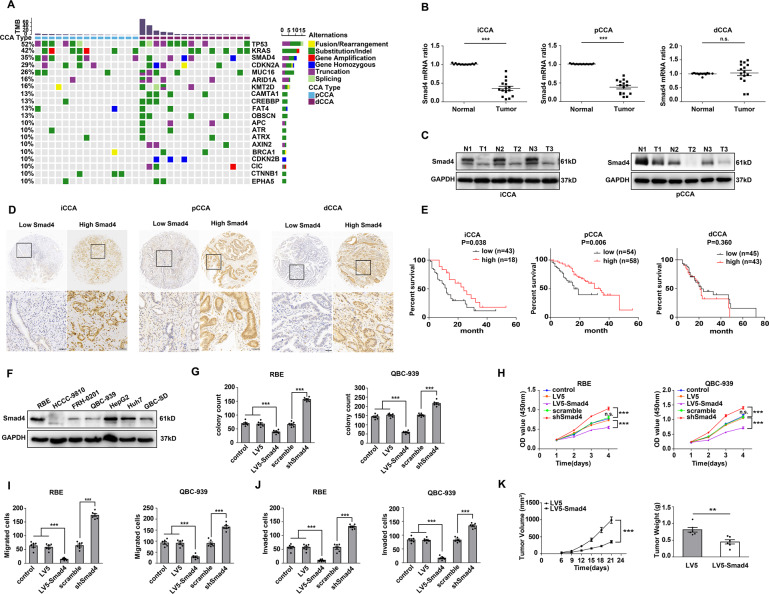
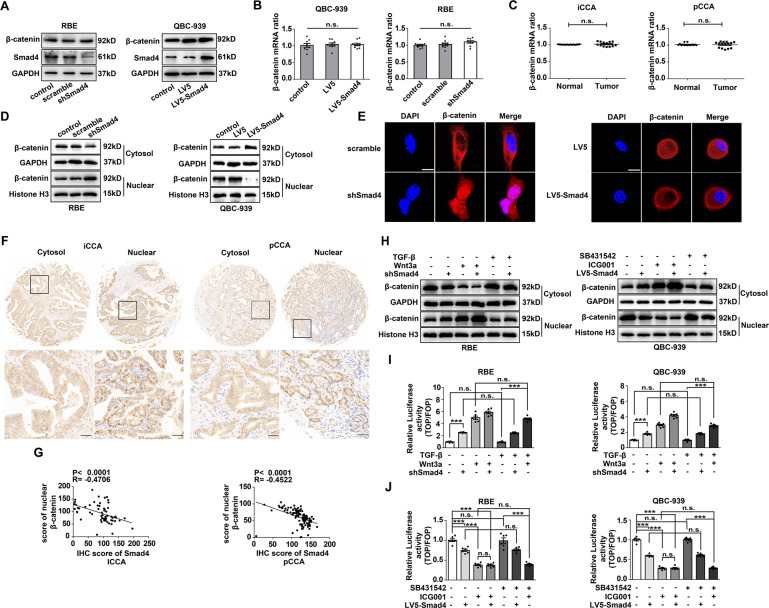
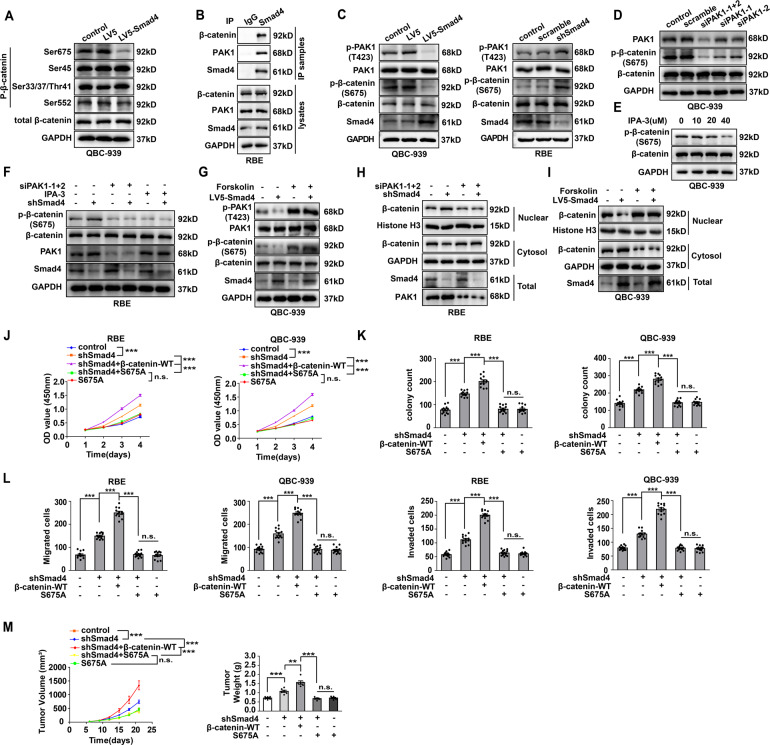
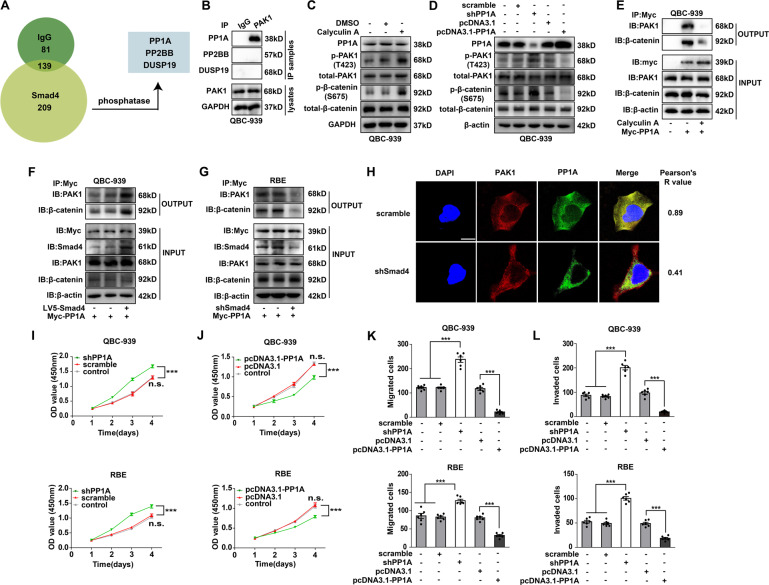
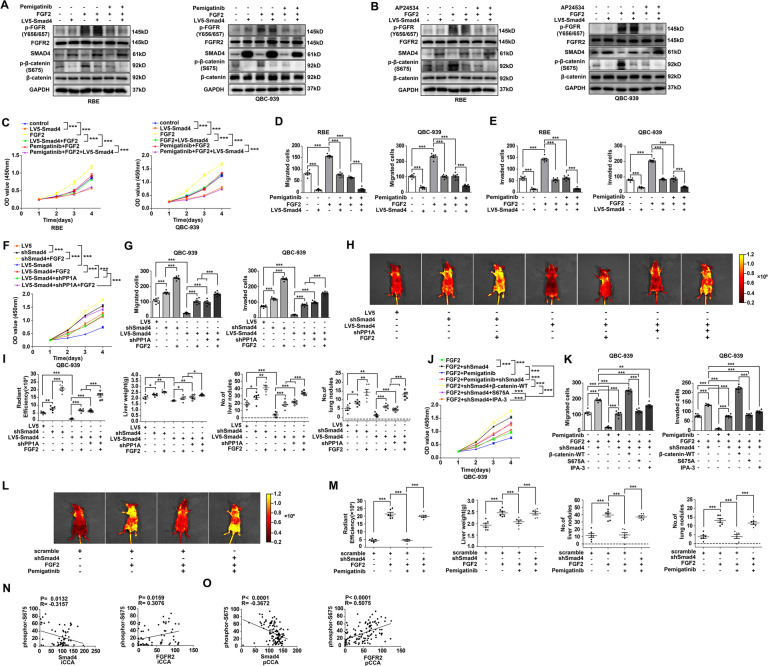
Cholangiocarcinoma (CCA), consisting of three subtypes-intrahepatic (iCCA), perihilar (pCCA), and distal (dCCA), is a highly aggressive cancer arising from the bile duct and has an extremely poor prognosis. Pemigatinib is the only FDA-approved targeted drug for CCA, and the CCA treatment options are substantially insufficient considering its poor prognosis and increasing morbidity. Here, we performed next-generation sequencing (NGS) of 15 pCCAs and 16 dCCAs and detected the expression of SMAD4, a frequently mutated gene, in 261 CCAs. By univariate and multivariate analyses, we identified Smad4 as a favorable prognostic biomarker in iCCA and pCCA. With in vitro and in vivo experiments, we demonstrated that Smad4 suppressed CCA proliferation, migration and invasion by inhibiting β-catenin-S675 phosphorylation and intranuclear translocation. We applied LC-MS/MS and multiple biochemical techniques and identified PP1A as the phosphatase in Smad4-mediated dephosphorylation of PAK1-T423, which is responsible for β-catenin-S675 phosphorylation. Moreover, we demonstrated that MYO18A is the PP1-interacting protein of PP1A for substrate recognition in CCA. MYO18A interacts with PP1A via its RVFFR motif and interacts with Smad4 via CC domain. Patients with coexpression of MYO18A and Smad4 have a more favorable prognosis than other patients. Smad4 enhances Pemigatinib efficiency, and Smad4 knockdown results in Pemigatinib resistance. In conclusion, coexpression of Smad4 and MYO18A is a favorable prognostic indicator for iCCA and pCCA. The Smad4-MYO18A-PP1A complex dephosphorylates PAK1-T423 and thus inhibits β-catenin-S675 phosphorylation and its intranuclear localization. Smad4 suppresses CCA proliferation, migration, invasion, and sensitivity to Pemigatinib by governing the phosphorylation and intracellular localization of β-catenin.
© 2021. The Author(s), under exclusive licence to ADMC Associazione Differenziamento e Morte Cellulare.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
