

Intestinal CD11b+ B Cells Ameliorate Colitis by Secreting Immunoglobulin A
- PMID: 34804004
- PMCID: PMC8595478
- DOI: 10.3389/fimmu.2021.697725
Intestinal CD11b+ B Cells Ameliorate Colitis by Secreting Immunoglobulin A
Abstract
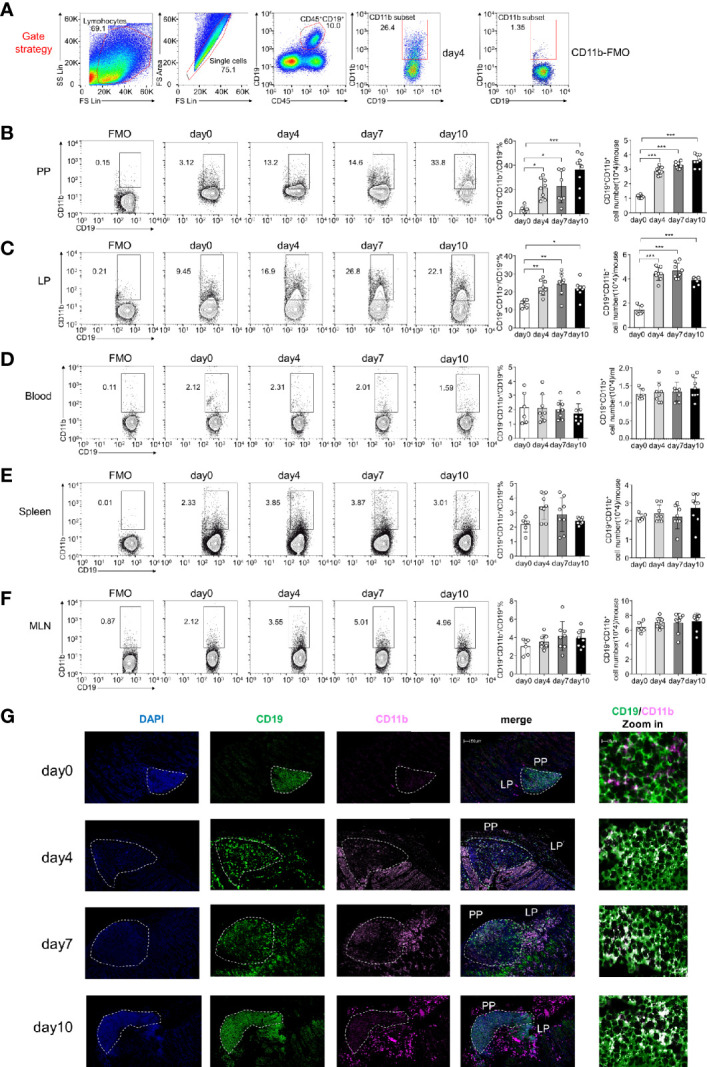
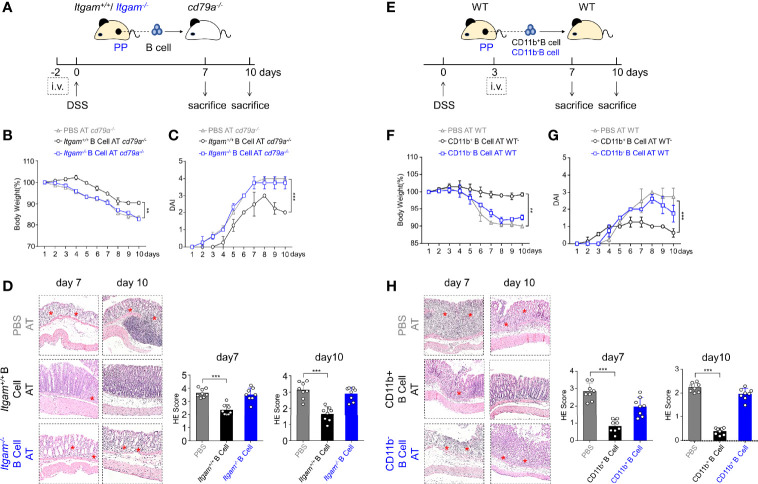
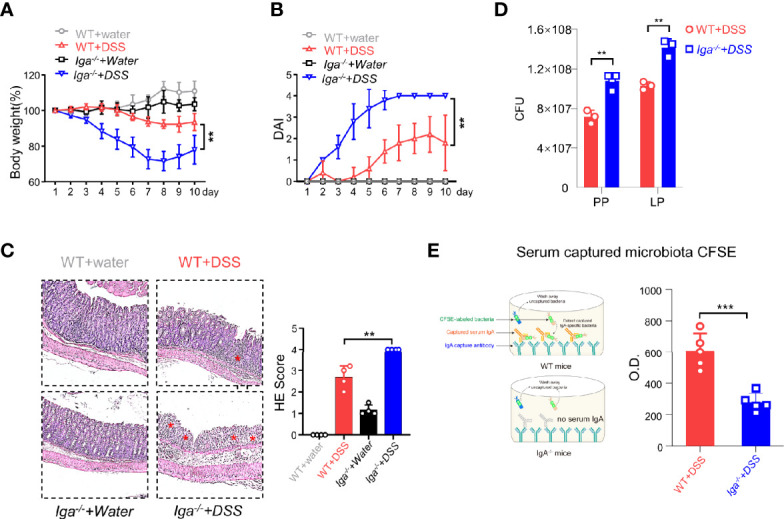
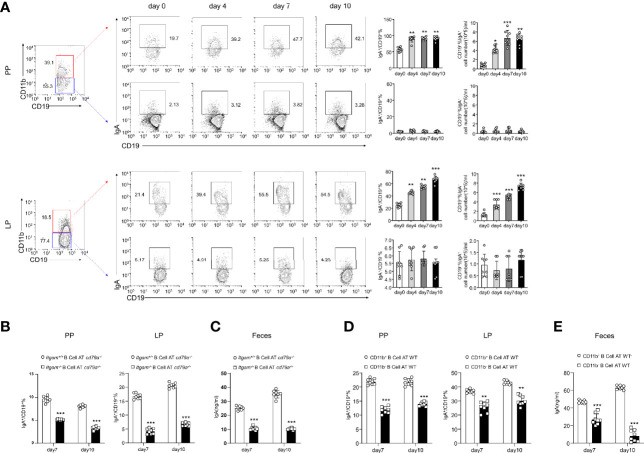
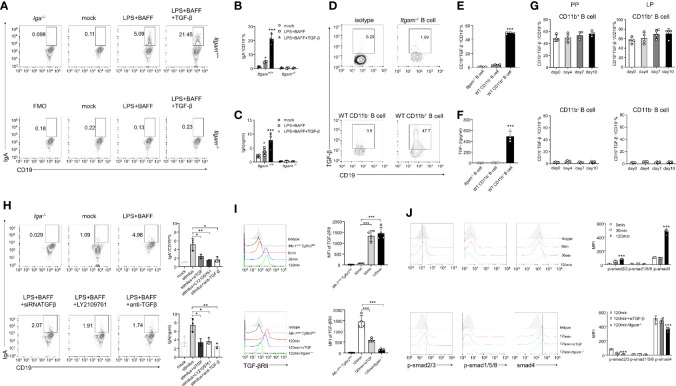
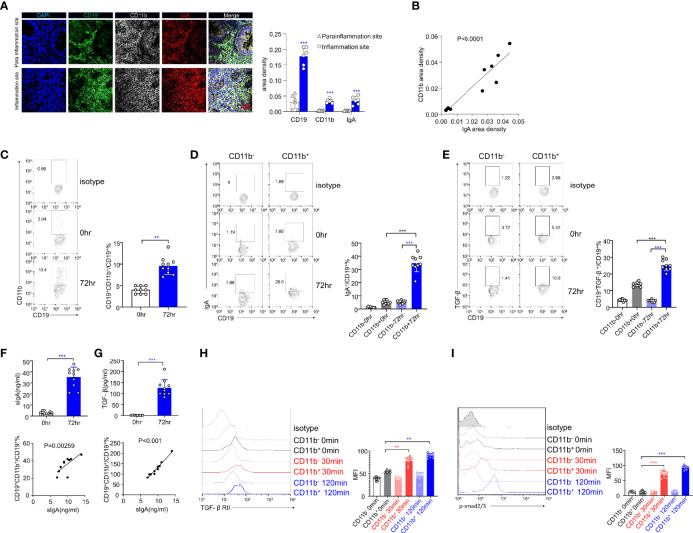
The intestinal mucosal immune environment requires multiple immune cells to maintain homeostasis. Although intestinal B cells are among the most important immune cells, little is known about the mechanism that they employ to regulate immune homeostasis. In this study, we found that CD11b+ B cells significantly accumulated in the gut lamina propria and Peyer's patches in dextran sulfate sodium-induced colitis mouse models and patients with ulcerative colitis. Adoptive transfer of CD11b+ B cells, but not CD11b-/- B cells, effectively ameliorated colitis and exhibited therapeutic effects. Furthermore, CD11b+ B cells were found to produce higher levels of IgA than CD11b- B cells. CD11b deficiency in B cells dampened IgA production, resulting in the loss of their ability to ameliorate colitis. Mechanistically, CD11b+ B cells expressed abundant TGF-β and TGF-β receptor II, as well as highly activate phosphorylated Smad2/3 signaling pathway, consequently promoting the class switch to IgA. Collectively, our findings demonstrate that CD11b+ B cells are essential intestinal suppressive immune cells and the primary source of intestinal IgA, which plays an indispensable role in maintaining intestinal homeostasis.
Keywords: CD11b+ B cells; DSS-induced colitis; IgA; Smad; TGF-β.
Copyright © 2021 Fu, Wang, Yu, Lin, Huang, Liu, Zhao, Lu, Xu, Liu, Liu, Wang and Chu.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Tseng J. Transfer of Lymphocytes of Peyer's Patches Between Immunoglobulin Allotype Congenic Mice: Repopulation of the IgA Plasma Cells in the Gut Lamina Propria. J Immunol (1981) 127(5):2039–43. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous