

Metabolic Enzyme Alterations and Astrocyte Dysfunction in a Murine Model of Alexander Disease With Severe Reactive Gliosis
- PMID: 34808356
- PMCID: PMC8717607
- DOI: 10.1016/j.mcpro.2021.100180
Metabolic Enzyme Alterations and Astrocyte Dysfunction in a Murine Model of Alexander Disease With Severe Reactive Gliosis
Abstract
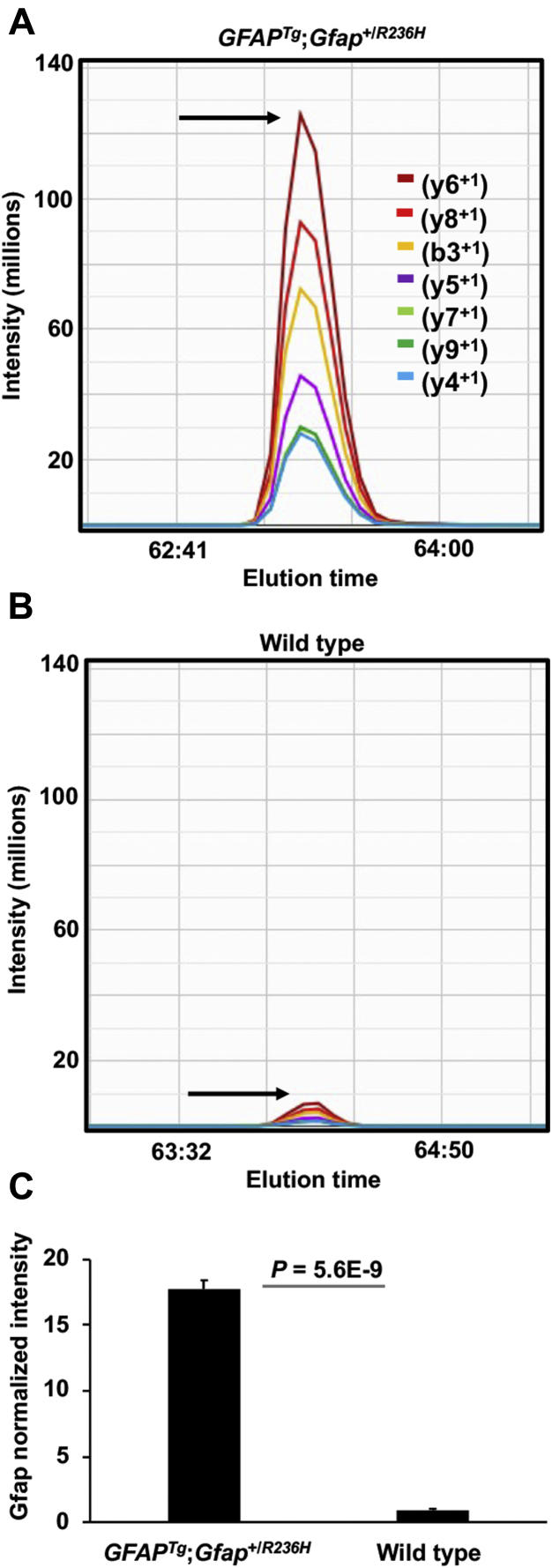
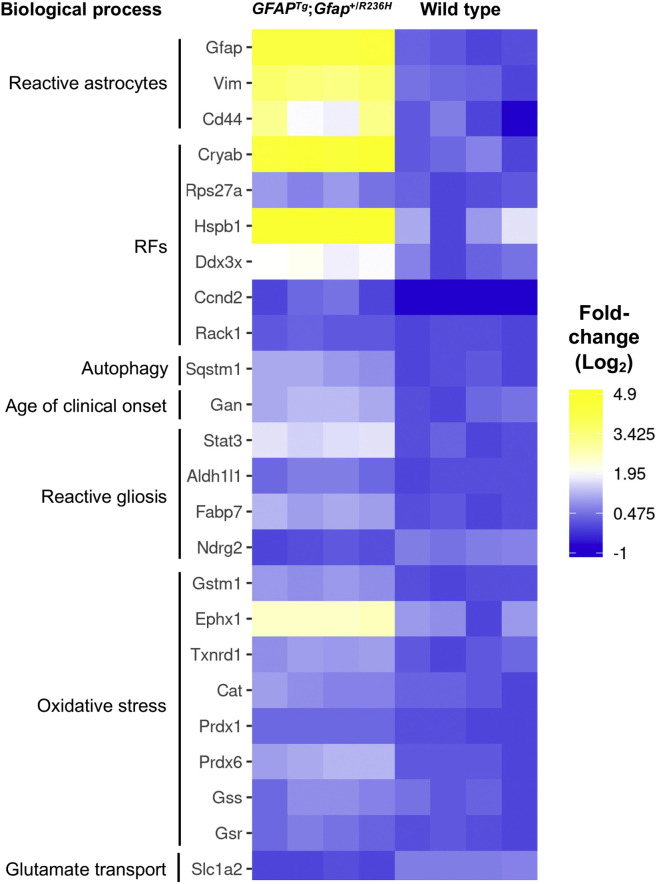
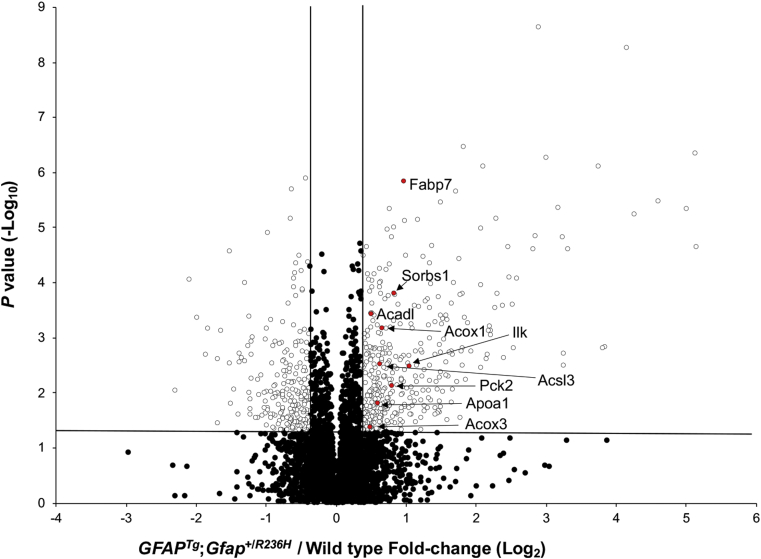
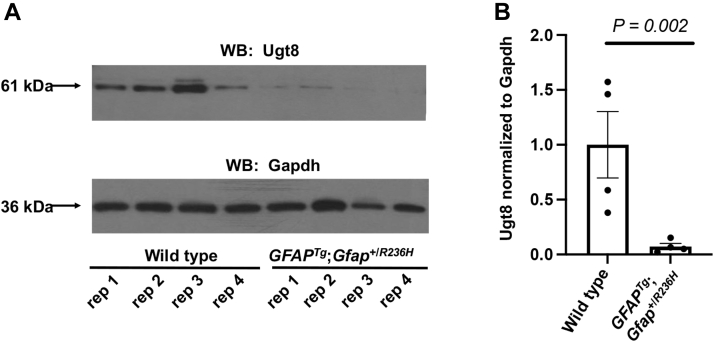
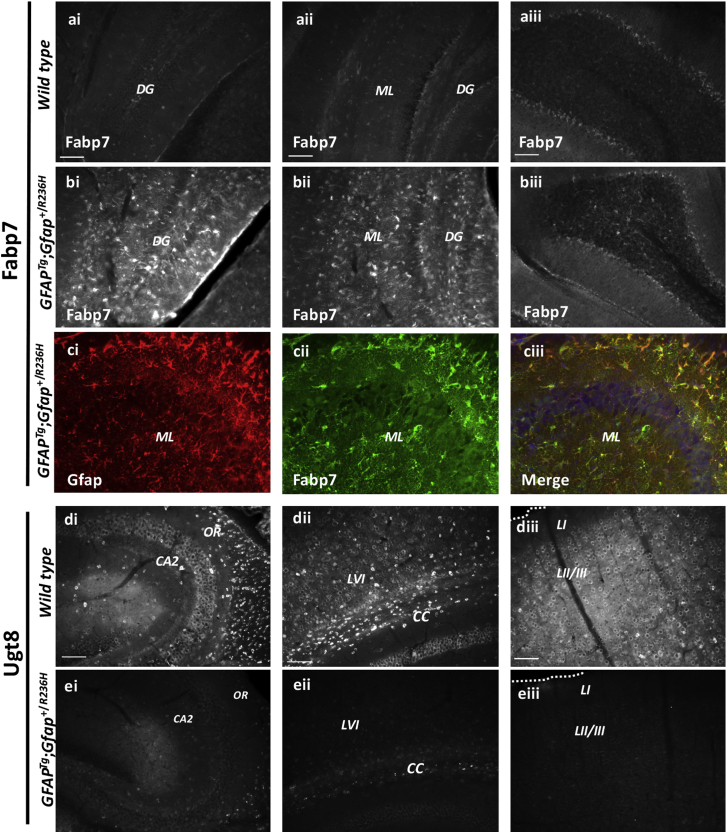
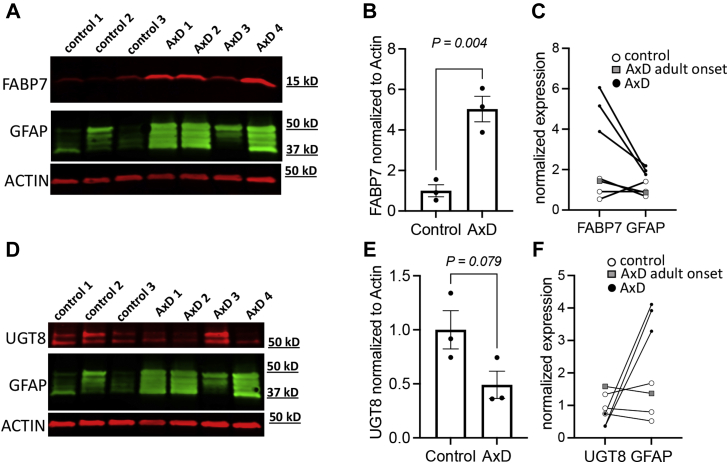
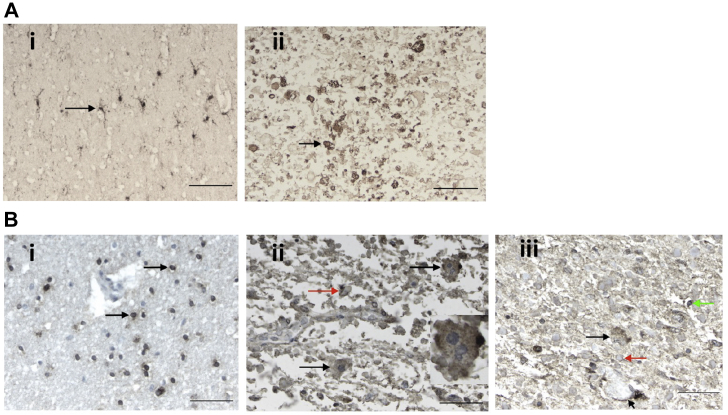
Alexander disease (AxD) is a rare and fatal neurodegenerative disorder caused by mutations in the gene encoding glial fibrillary acidic protein (GFAP). In this report, a mouse model of AxD (GFAPTg;Gfap+/R236H) was analyzed that contains a heterozygous R236H point mutation in murine Gfap as well as a transgene with a GFAP promoter to overexpress human GFAP. Using label-free quantitative proteomic comparisons of brain tissue from GFAPTg;Gfap+/R236H versus wild-type mice confirmed upregulation of the glutathione metabolism pathway and indicated proteins were elevated in the peroxisome proliferator-activated receptor (PPAR) signaling pathway, which had not been reported previously in AxD. Relative protein-level differences were confirmed by a targeted proteomics assay, including proteins related to astrocytes and oligodendrocytes. Of particular interest was the decreased level of the oligodendrocyte protein, 2-hydroxyacylsphingosine 1-beta-galactosyltransferase (Ugt8), since Ugt8-deficient mice exhibit a phenotype similar to GFAPTg;Gfap+/R236H mice (e.g., tremors, ataxia, hind-limb paralysis). In addition, decreased levels of myelin-associated proteins were found in the GFAPTg;Gfap+/R236H mice, consistent with the role of Ugt8 in myelin synthesis. Fabp7 upregulation in GFAPTg;Gfap+/R236H mice was also selected for further investigation due to its uncharacterized association to AxD, critical function in astrocyte proliferation, and functional ability to inhibit the anti-inflammatory PPAR signaling pathway in models of amyotrophic lateral sclerosis (ALS). Within Gfap+ astrocytes, Fabp7 was markedly increased in the hippocampus, a brain region subjected to extensive pathology and chronic reactive gliosis in GFAPTg;Gfap+/R236H mice. Last, to determine whether the findings in GFAPTg;Gfap+/R236H mice are present in the human condition, AxD patient and control samples were analyzed by Western blot, which indicated that Type I AxD patients have a significant fourfold upregulation of FABP7. However, immunohistochemistry analysis showed that UGT8 accumulates in AxD patient subpial brain regions where abundant amounts of Rosenthal fibers are located, which was not observed in the GFAPTg;Gfap+/R236H mice.
Keywords: Alexander disease; Fabp7; Ugt8; astrocytes; reactive gliosis.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors have no conflicts of interest in the study.
Figures
References
-
- Zhang Y., Sloan S.A., Clarke L.E., Caneda C., Plaza C.A., Blumenthal P.D., Vogel H., Steinberg G.K., Edwards M.S., Li G., Duncan J.A., 3rd, Cheshier S.H., Shuer L.M., Chang E.F., Grant G.A., et al. Purification and characterization of progenitor and mature human astrocytes reveals transcriptional and functional differences with mouse. Neuron. 2016;89:37–53. - PMC - PubMed
-
- Rothstein J.D., Dykes-Hoberg M., Pardo C.A., Bristol L.A., Jin L., Kuncl R.W., Kanai Y., Hediger M.A., Wang Y., Schielke J.P., Welty D.F. Knockout of glutamate transporters reveals a major role for astroglial transport in excitotoxicity and clearance of glutamate. Neuron. 1996;16:675–686. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
