

Microbiota links to neural dynamics supporting threat processing
- PMID: 34811847
- PMCID: PMC8720184
- DOI: 10.1002/hbm.25682
Microbiota links to neural dynamics supporting threat processing
Abstract
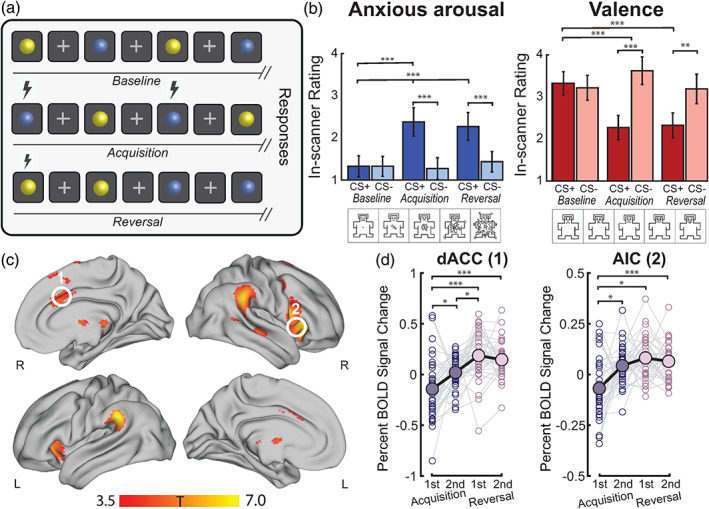
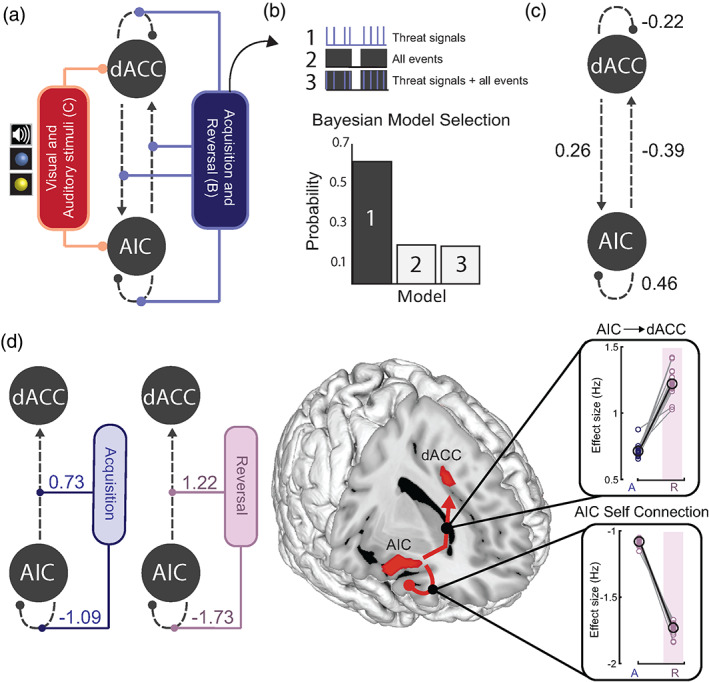
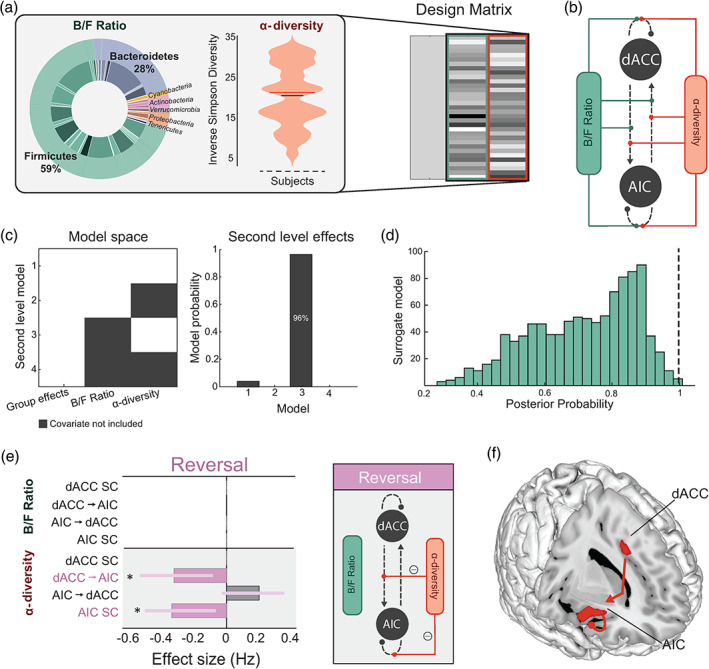
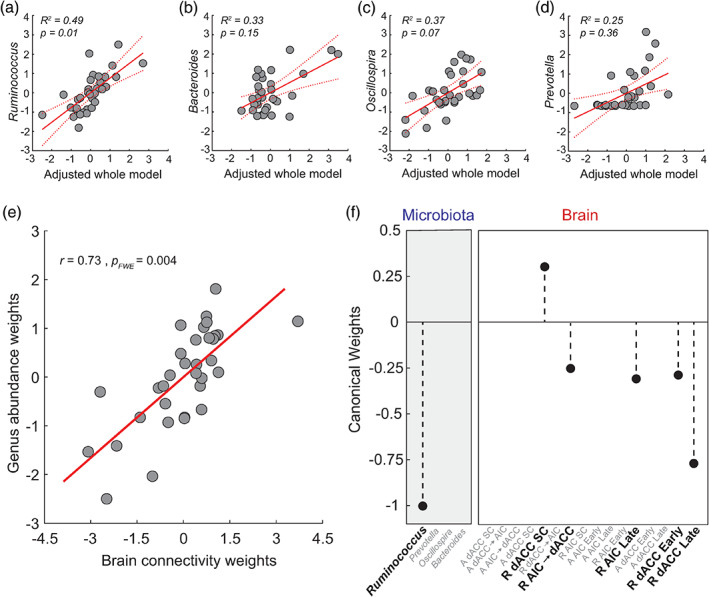
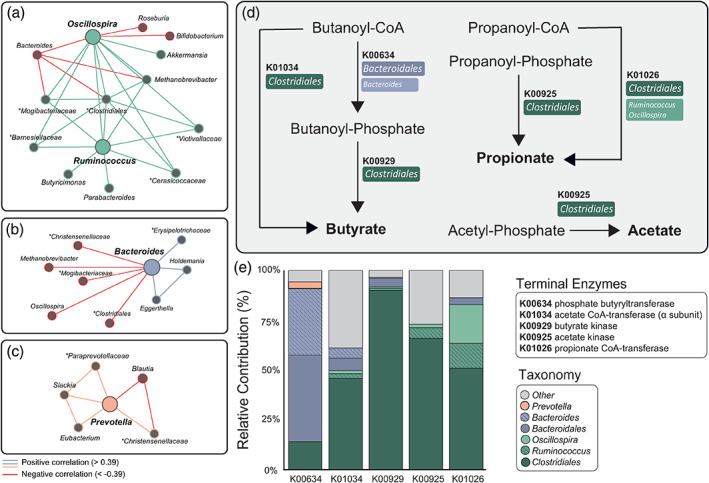
There is growing recognition that the composition of the gut microbiota influences behaviour, including responses to threat. The cognitive-interoceptive appraisal of threat-related stimuli relies on dynamic neural computations between the anterior insular (AIC) and the dorsal anterior cingulate (dACC) cortices. If, to what extent, and how microbial consortia influence the activity of this cortical threat processing circuitry is unclear. We addressed this question by combining a threat processing task, neuroimaging, 16S rRNA profiling and computational modelling in healthy participants. Results showed interactions between high-level ecological indices with threat-related AIC-dACC neural dynamics. At finer taxonomic resolutions, the abundance of Ruminococcus was differentially linked to connectivity between, and activity within the AIC and dACC during threat updating. Functional inference analysis provides a strong rationale to motivate future investigations of microbiota-derived metabolites in the observed relationship with threat-related brain processes.
Keywords: anterior insula; dorsal anterior cingulate; gut-brain axis; microbiota; neuroimaging; threat processing.
© 2021 The Authors. Human Brain Mapping published by Wiley Periodicals LLC.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Aatsinki, A.‐K. , Lahti, L. , Uusitupa, H.‐M. , Munukka, E. , Keskitalo, A. , Nolvi, S. , … Karlsson, L. (2019). Gut microbiota composition is associated with temperament traits in infants. Brain, Behavior, and Immunity, 80, 849–858. - PubMed
-
- Allman, J. M. , Tetreault, N. A. , Hakeem, A. Y. , Manaye, K. F. , Semendeferi, K. , Erwin, J. M. , … Hof, P. R. (2010). The von Economo neurons in frontoinsular and anterior cingulate cortex in great apes and humans. Brain Structure and Function, 214, 495–517. - PubMed
-
- Azzalini, D. , Rebollo, I. , & Tallon‐Baudry, C. (2019). Visceral signals shape brain dynamics and cognition. Trends in Cognitive Sciences, 23, 488–509. - PubMed
-
- Bastian, M. , Heymann, S. , Jacomy, M. , 2009. Gephi: An open source software for exploring and manipulating networks. Third International AAAI Conference on Weblogs and Social Media.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
