

Spatial and Feature-selective Attention Have Distinct, Interacting Effects on Population-level Tuning
- PMID: 34813647
- PMCID: PMC7613071
- DOI: 10.1162/jocn_a_01796
Spatial and Feature-selective Attention Have Distinct, Interacting Effects on Population-level Tuning
Abstract
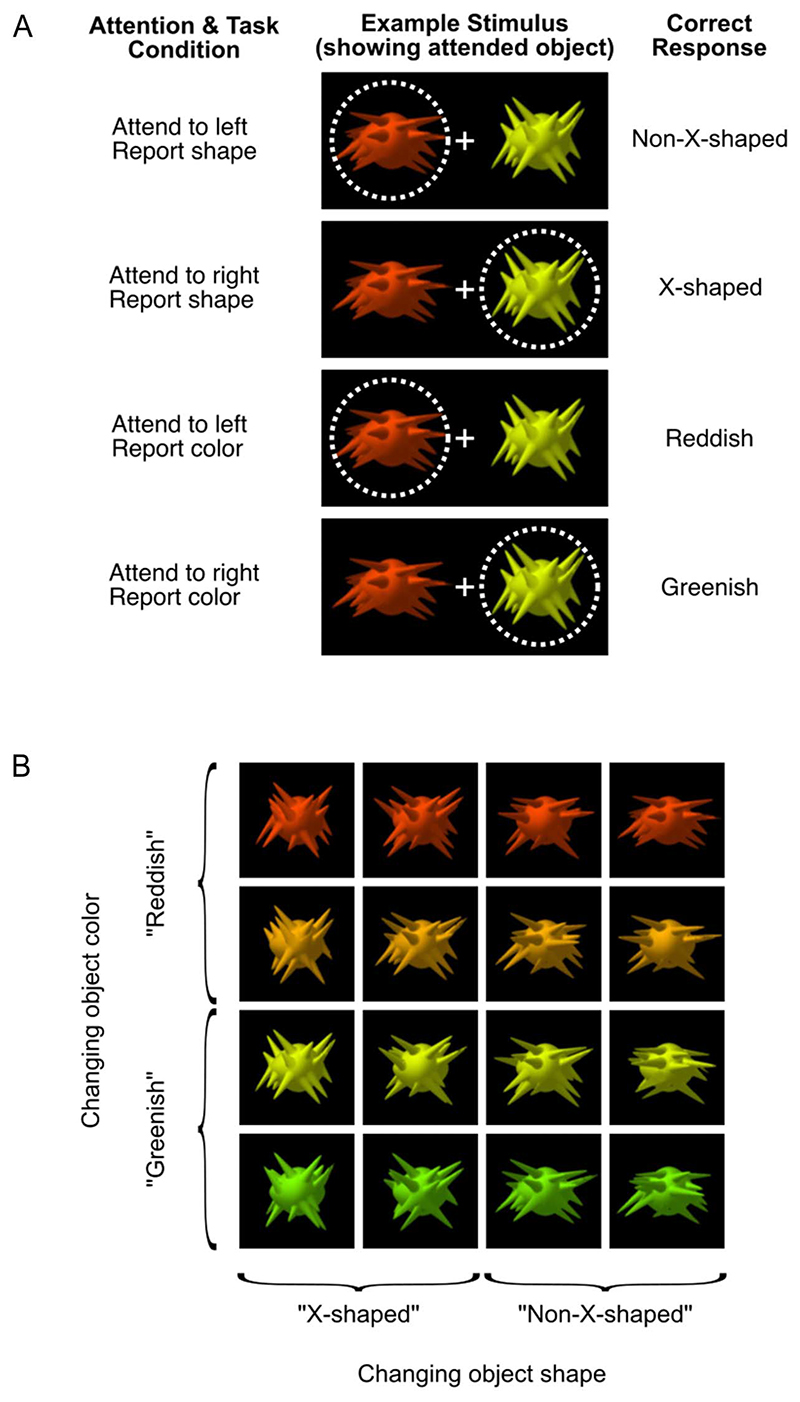
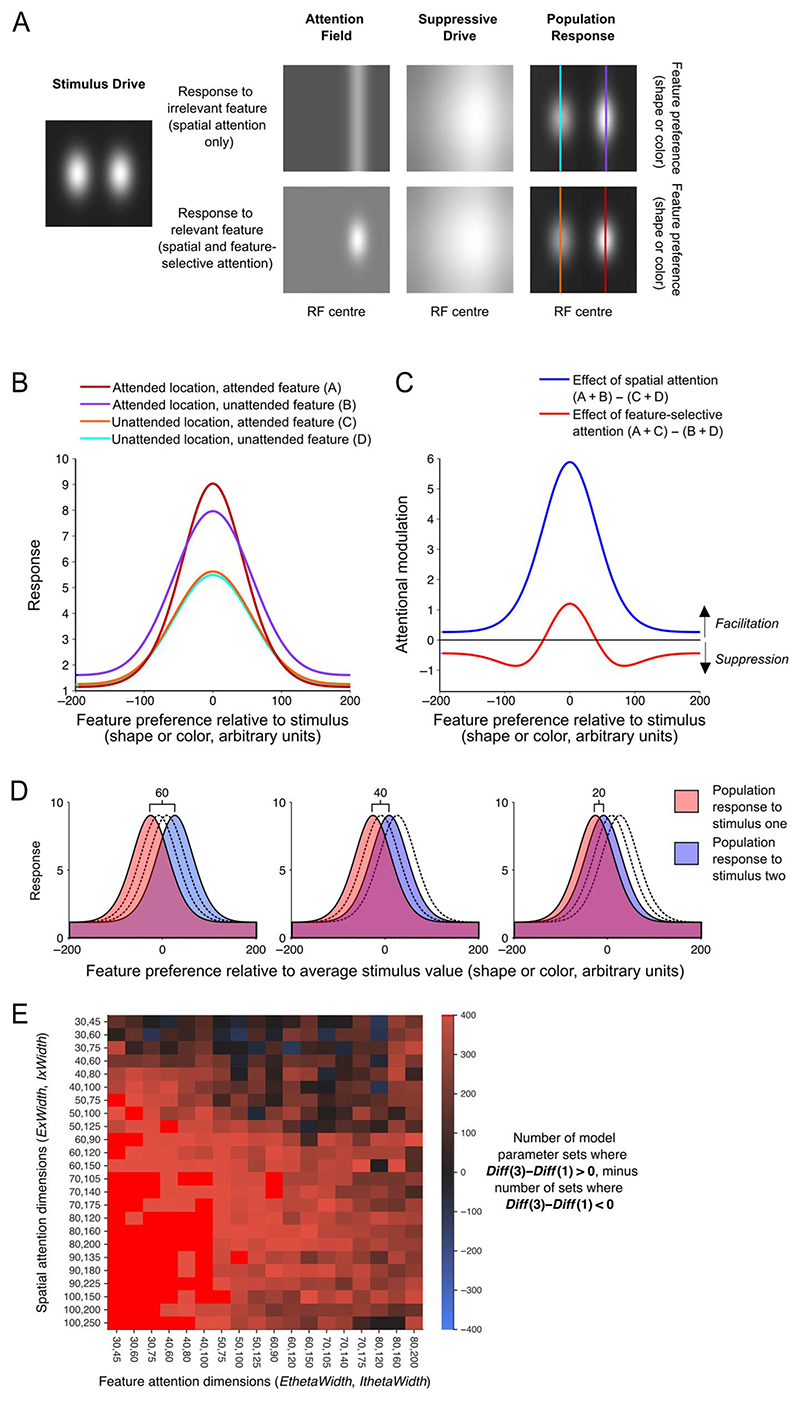
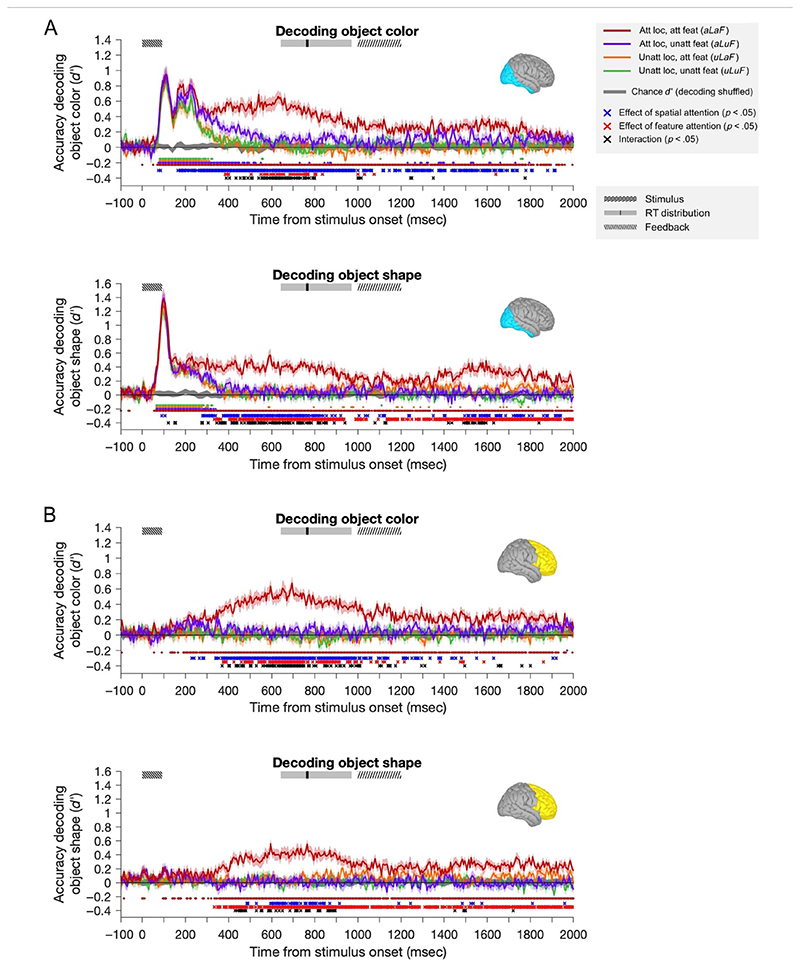
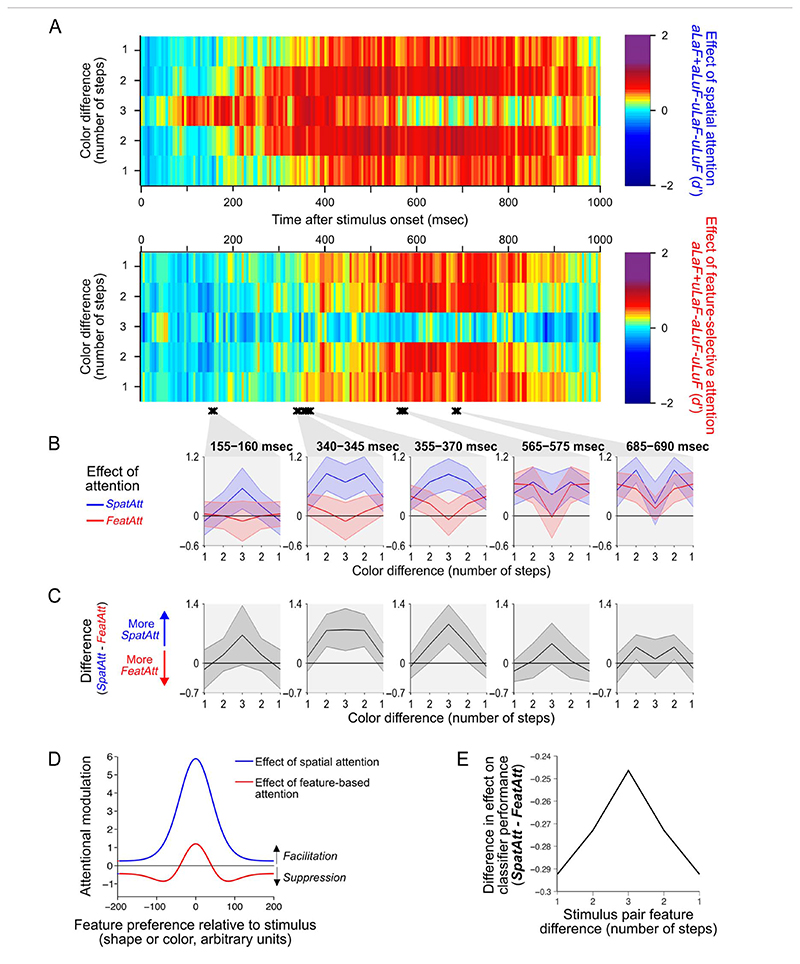
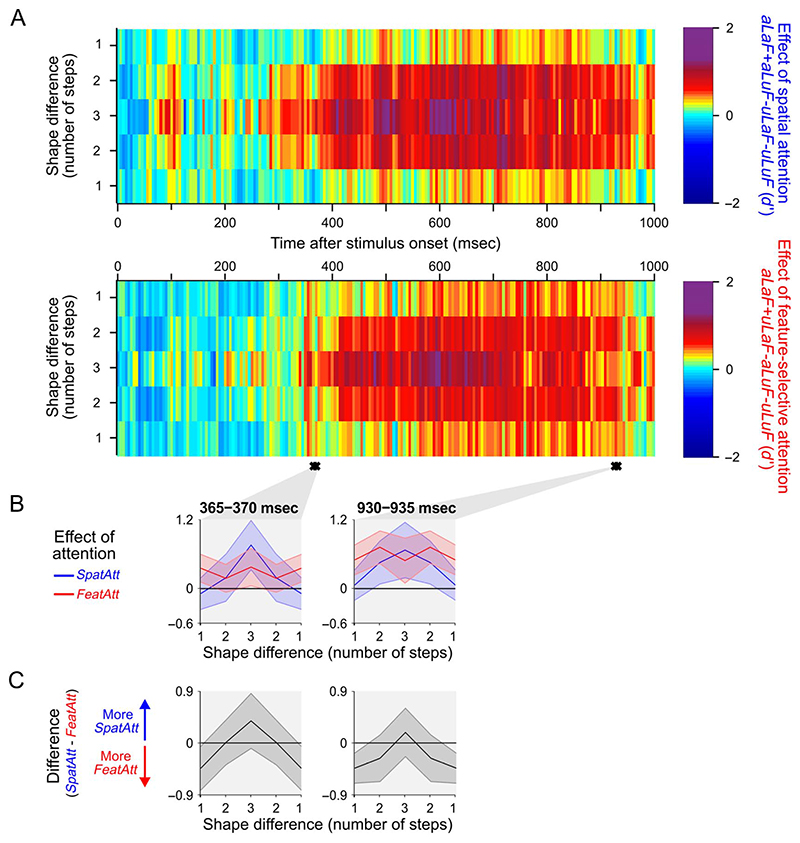
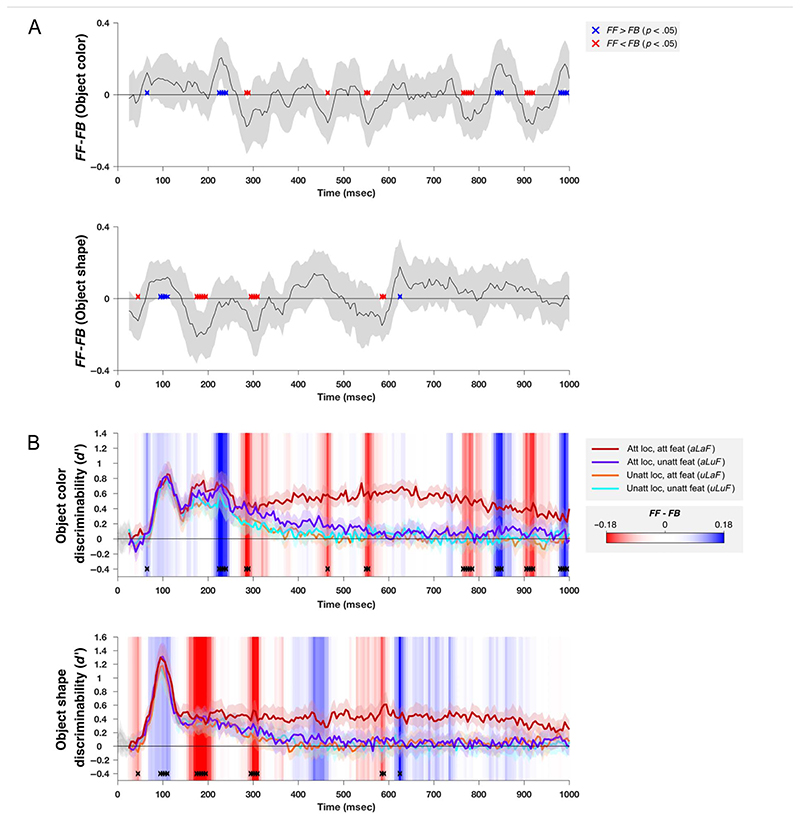
Attention can be deployed in different ways: When searching for a taxi in New York City, we can decide where to attend (e.g., to the street) and what to attend to (e.g., yellow cars). Although we use the same word to describe both processes, nonhuman primate data suggest that these produce distinct effects on neural tuning. This has been challenging to assess in humans, but here we used an opportunity afforded by multivariate decoding of MEG data. We found that attending to an object at a particular location and attending to a particular object feature produced effects that interacted multiplicatively. The two types of attention induced distinct patterns of enhancement in occipital cortex, with feature-selective attention producing relatively more enhancement of small feature differences and spatial attention producing relatively larger effects for larger feature differences. An information flow analysis further showed that stimulus representations in occipital cortex were Granger-caused by coding in frontal cortices earlier in time and that the timing of this feedback matched the onset of attention effects. The data suggest that spatial and feature-selective attention rely on distinct neural mechanisms that arise from frontal-occipital information exchange, interacting multiplicatively to selectively enhance task-relevant information.
© 2021 the Massachusetts Institute of Technology. Published under a Creative Commons Attribution 4.0 International (CC BY 4.0) license.
Figures
References
-
- Benjamini Y, Hochberg Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. Journal of the Royal Statistical Society, Series B: Methodological. 1995;57:289–300. doi: 10.1111/j.2517-6161.1995.tb02031.x. - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources