

Noncanonical protease-activated receptor 1 regulates lymphatic differentiation in zebrafish
- PMID: 34816109
- PMCID: PMC8593614
- DOI: 10.1016/j.isci.2021.103386
Noncanonical protease-activated receptor 1 regulates lymphatic differentiation in zebrafish
Abstract
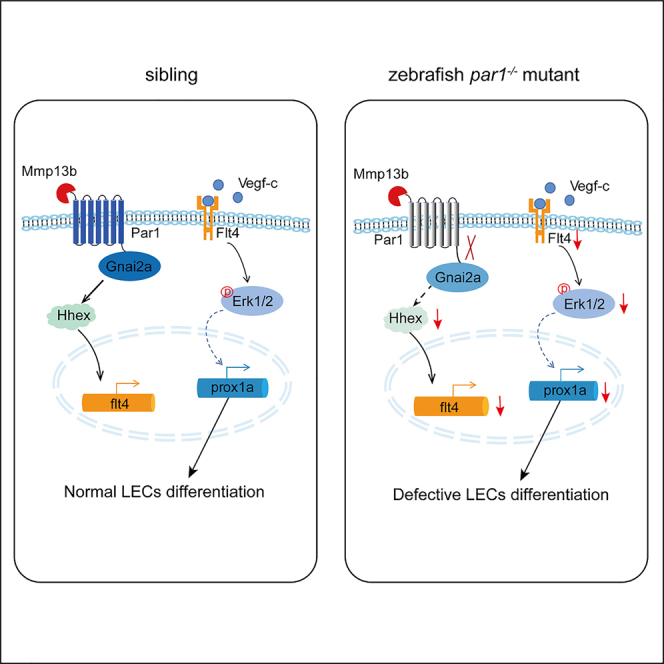
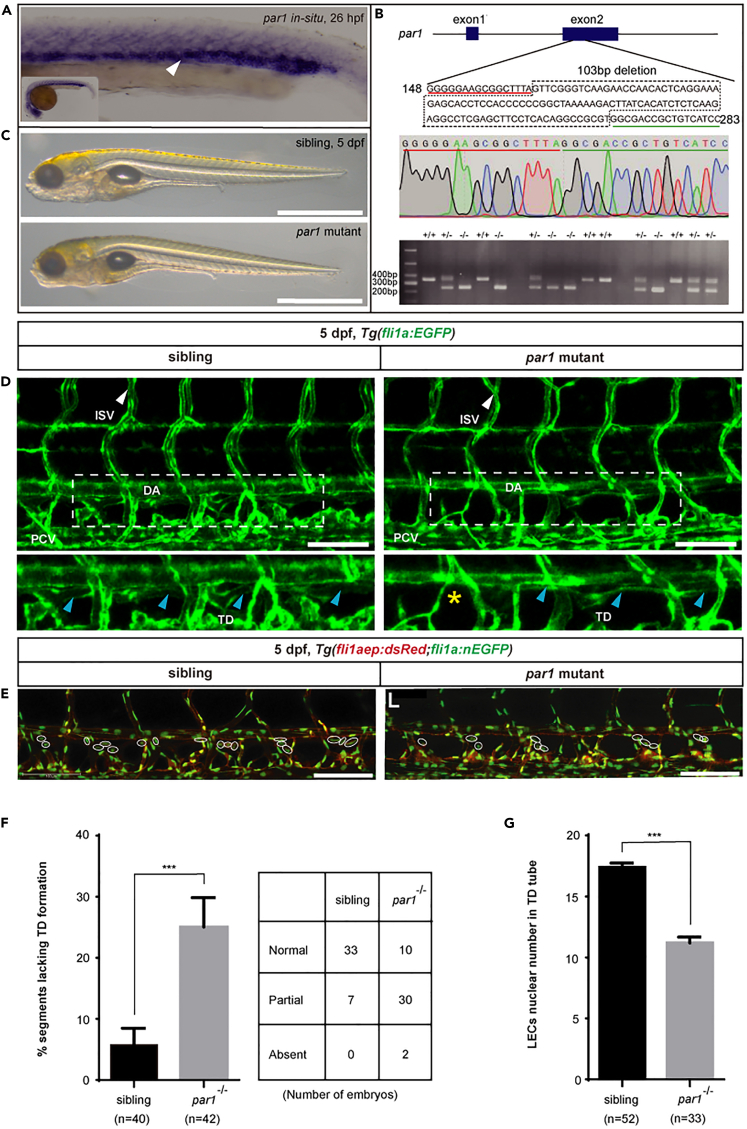
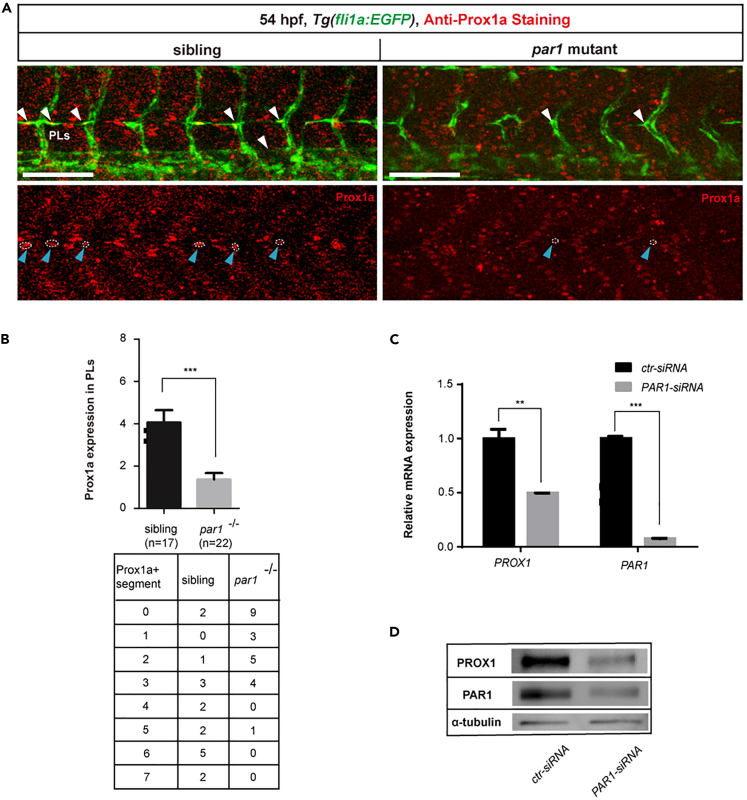
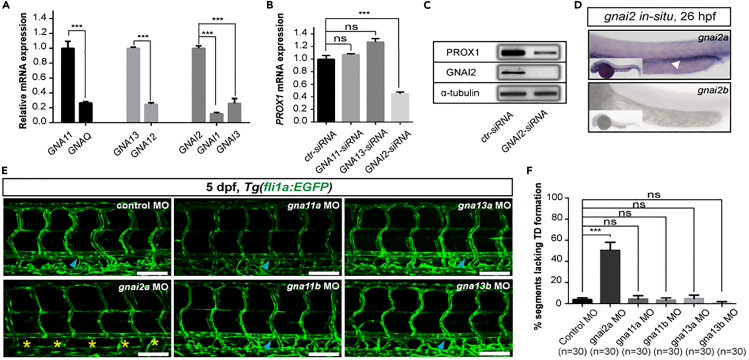
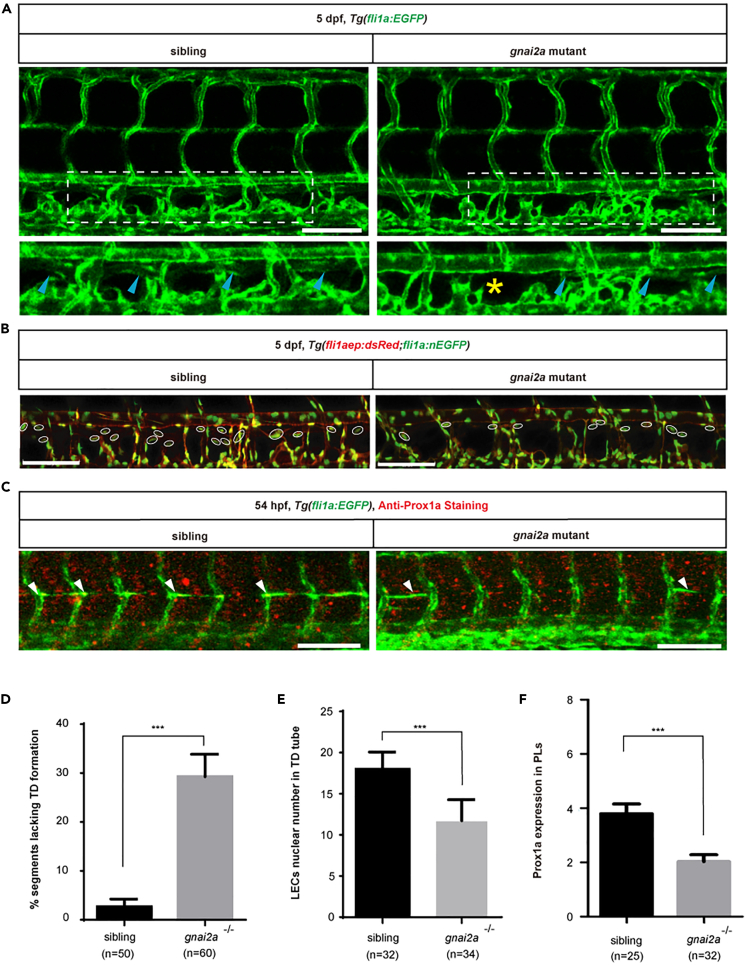
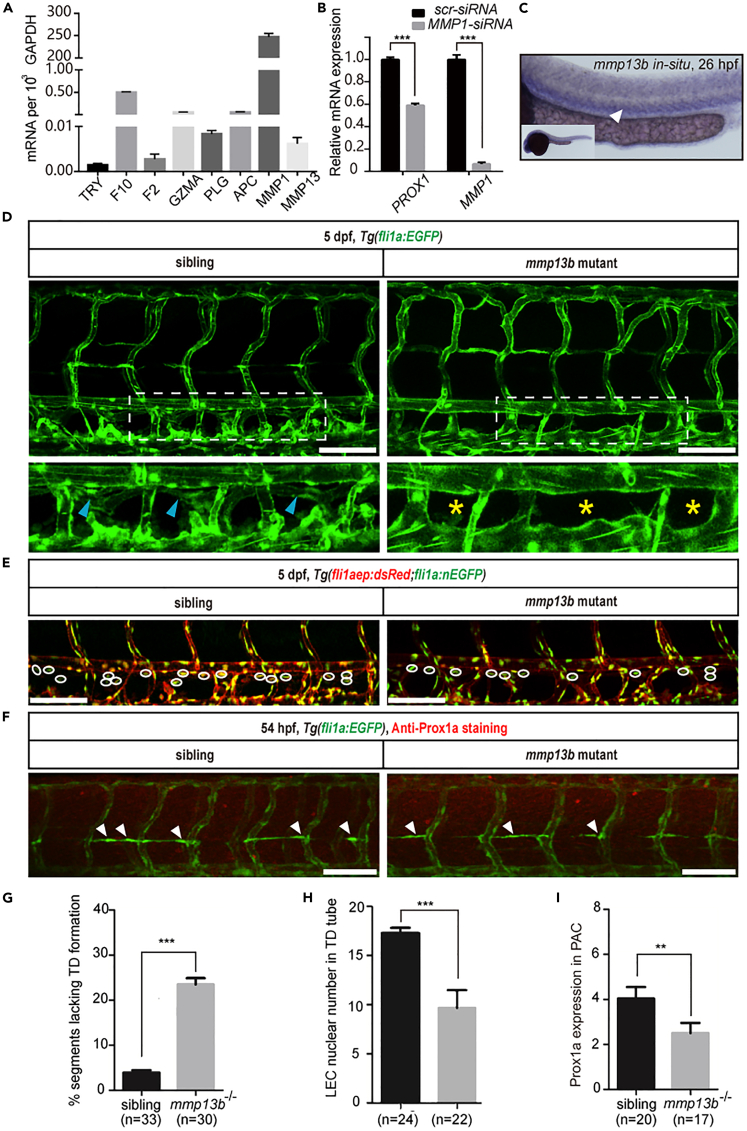
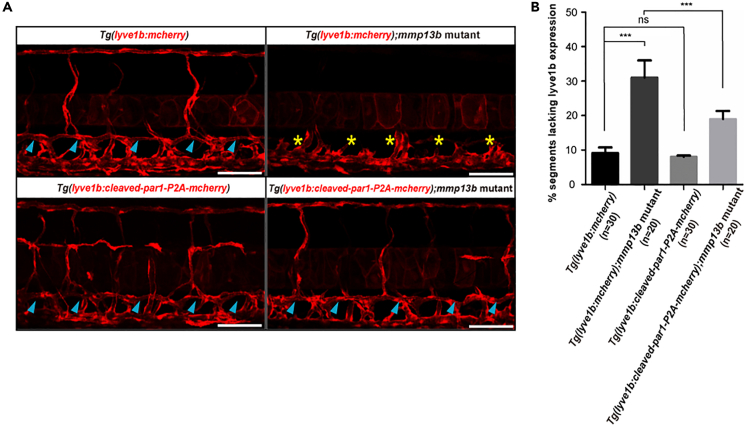
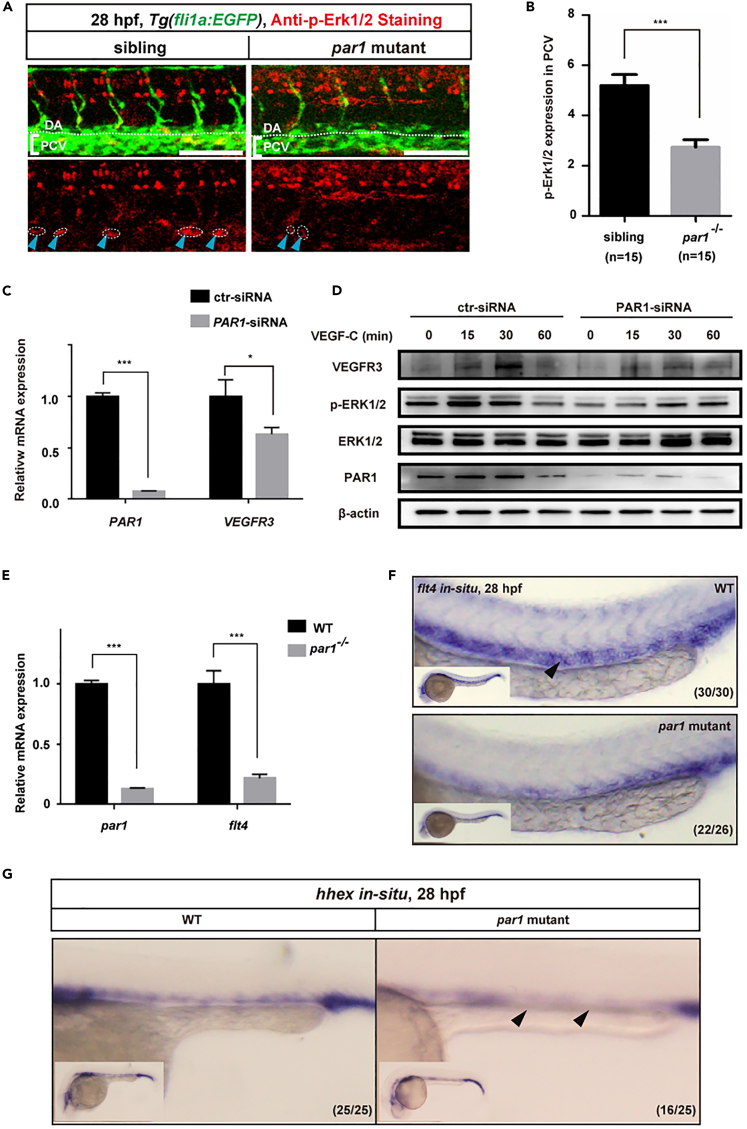
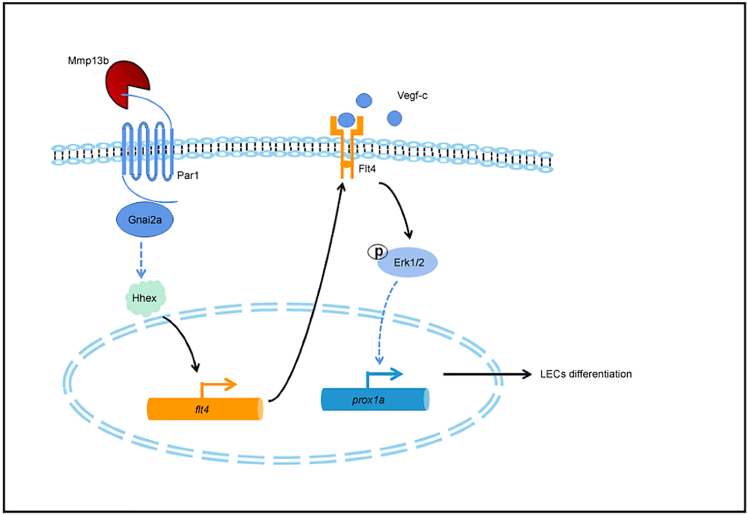
The differentiation of lymphatic progenitors is a crucial step in lymphangiogenesis. However, its underlying mechanism remains unclear. Here, we found that noncanonical protease-activated receptor 1 (par1) regulates the differentiation of lymphatic progenitors in zebrafish embryos. Loss of par1 function impaired lymphatic differentiation by downregulating prox1a expression in parachordal lymphangioblasts and caused compromised thoracic duct formation in zebrafish. Meanwhile, the G protein gnai2a, a par1 downstream effector, was selectively required for lymphatic development in zebrafish, and its mutation mimicked the lymphatic phenotype observed in par1 mutants. Interestingly, mmp13, but not thrombin, was required for lymphatic development in zebrafish. Furthermore, analyses of genetic interactions confirmed that mmp13b serves as a par1 upstream protease to regulate lymphatic development in zebrafish embryos. Mechanistically, par1 promotes flt4 expression and phospho-Erk1/2 activity in the posterior cardinal vein. Taken together, our findings highlight a function of par1 in the regulation of lymphatic differentiation in zebrafish embryos.
Keywords: Biological sciences; Immunology; Molecular biology.
© 2021 The Author(s).
Conflict of interest statement
The authors declare that there is no conflict of interest.
Figures
References
-
- Austin K.M., Nguyen N., Javid G., Covic L., Kuliopulos A. Noncanonical matrix metalloprotease-1-protease-activated receptor-1 signaling triggers vascular smooth muscle cell dedifferentiation and arterial stenosis. J. Biol. Chem. 2013;288:23105–23115. doi: 10.1074/jbc.M113.467019. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
