

Proteomic dissection of large extracellular vesicle surfaceome unravels interactive surface platform
- PMID: 34817906
- PMCID: PMC8612312
- DOI: 10.1002/jev2.12164
Proteomic dissection of large extracellular vesicle surfaceome unravels interactive surface platform
Abstract
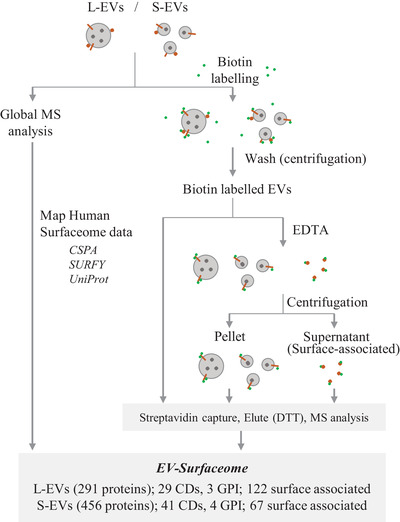
The extracellular vesicle (EV) surface proteome (surfaceome) acts as a fundamental signalling gateway by bridging intra- and extracellular signalling networks, dictates EVs' capacity to communicate and interact with their environment, and is a source of potential disease biomarkers and therapeutic targets. However, our understanding of surface protein composition of large EVs (L-EVs, 100-800 nm, mean 310 nm, ATP5F1A, ATP5F1B, DHX9, GOT2, HSPA5, HSPD1, MDH2, STOML2), a major EV-subtype that are distinct from small EVs (S-EVs, 30-150 nm, mean 110 nm, CD44, CD63, CD81, CD82, CD9, PDCD6IP, SDCBP, TSG101) remains limited. Using a membrane impermeant derivative of biotin to capture surface proteins coupled to mass spectrometry analysis, we show that out of 4143 proteins identified in density-gradient purified L-EVs (1.07-1.11 g/mL, from multiple cancer cell lines), 961 proteins are surface accessible. The surface molecular diversity of L-EVs include (i) bona fide plasma membrane anchored proteins (cluster of differentiation, transporters, receptors and GPI anchored proteins implicated in cell-cell and cell-ECM interactions); and (ii) membrane surface-associated proteins (that are released by divalent ion chelator EDTA) implicated in actin cytoskeleton regulation, junction organization, glycolysis and platelet activation. Ligand-receptor analysis of L-EV surfaceome (e.g., ITGAV/ITGB1) uncovered interactome spanning 172 experimentally verified cognate binding partners (e.g., ANGPTL3, PLG, and VTN) with highest tissue enrichment for liver. Assessment of biotin inaccessible L-EV proteome revealed enrichment for proteins belonging to COPI/II-coated ER/Golgi-derived vesicles and mitochondria. Additionally, despite common surface proteins identified in L-EVs and S-EVs, our data reveals surfaceome heterogeneity between the two EV-subtype. Collectively, our study provides critical insights into diverse proteins operating at the interactive platform of L-EVs and molecular leads for future studies seeking to decipher L-EV heterogeneity and function.
Keywords: extracellular vesicles; mass spectrometry-based proteomics; surface proteins; surfaceome; vesicle heterogeneity.
© 2021 The Authors. Journal of Extracellular Vesicles published by Wiley Periodicals, LLC on behalf of the International Society for Extracellular Vesicles.
Figures





References
-
- Al‐Nedawi, K. , Meehan, B. , Micallef, J. , Lhotak, V. , May, L. , Guha, A. , & Rak, J. (2008). Intercellular transfer of the oncogenic receptor EGFRvIII by microvesicles derived from tumour cells. Nature Cell Biology, 10, 619–624. - PubMed
-
- Bausch‐Fluck, D. , Hofmann, A. , Bock, T. , Frei, A. P. , Cerciello, F. , Jacobs, A. , Moest, H. , Omasits, U. , Gundry, R. L. , Yoon, C. , Schiess, R. , Schmidt, A. , Mirkowska, P. , Härtlová, A. , Van Eyk, J. E. , Bourquin, J.‐P. , Aebersold, R. , Boheler, K. R. , Zandstra, P. , & Wollscheid, B. (2015). A mass spectrometric‐derived cell surface protein atlas. Plos One, 10, e0121314. - PMC - PubMed
-
- Bebawy, M. , Combes, V. , Lee, E. , Jaiswal, R. , Gong, J. , Bonhoure, A. , Grau, G. E. R. (2009). Membrane microparticles mediate transfer of P‐glycoprotein to drug sensitive cancer cells. Leukemia, 23, 1643–1649. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
