

TM6SF2/PNPLA3/MBOAT7 Loss-of-Function Genetic Variants Impact on NAFLD Development and Progression Both in Patients and in In Vitro Models
- PMID: 34823063
- PMCID: PMC8783129
- DOI: 10.1016/j.jcmgh.2021.11.007
TM6SF2/PNPLA3/MBOAT7 Loss-of-Function Genetic Variants Impact on NAFLD Development and Progression Both in Patients and in In Vitro Models
Abstract
Background & aims: The I148M Patatin-like Phospholipase Domain-containing 3 (PNPLA3), the rs641738 in the Membrane bound O-acyltransferase domain containing 7-transmembrane channel-like 4 (MBOAT7-TMC4) locus, and the E167K Transmembrane 6 Superfamily Member 2 (TM6SF2) polymorphisms represent the main predisposing factors to nonalcoholic fatty liver disease (NAFLD) development and progression. We previously generated a full knockout of MBOAT7 in HepG2 cells (MBOAT7-/-), homozygous for I148M PNPLA3. Therefore, we aimed to investigate the synergic impact of the 3 at-risk variants on liver injury and hepatocellular carcinoma (HCC) in a large cohort of NAFLD patients, and create in vitro models of genetic NAFLD by silencing TM6SF2 in both HepG2 and MBOAT7-/- cells.
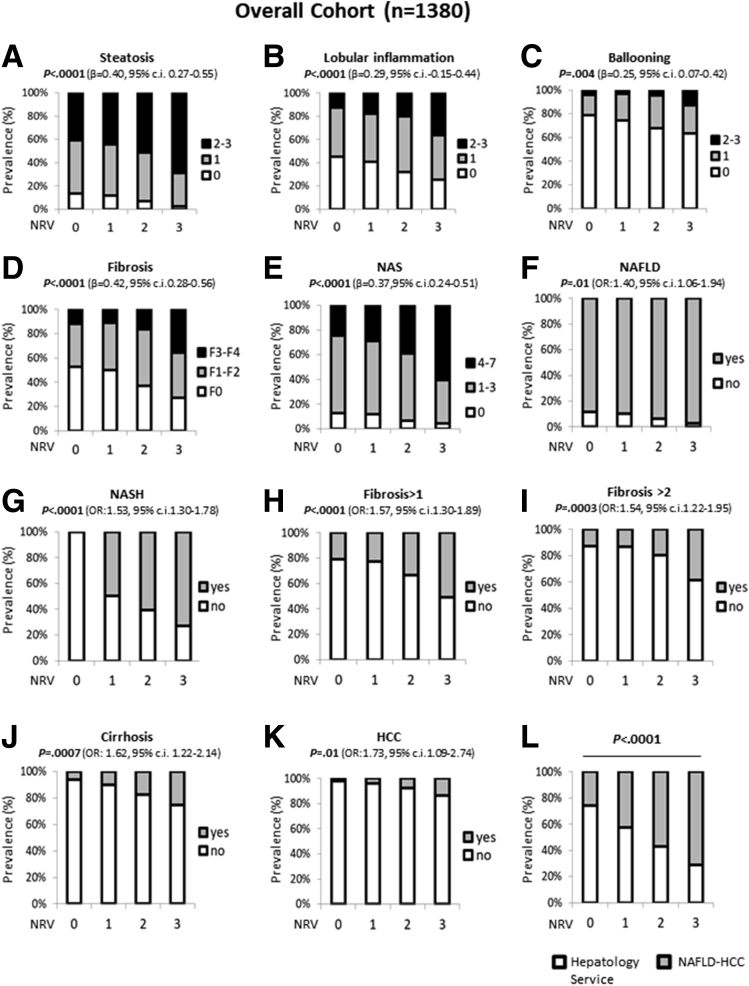
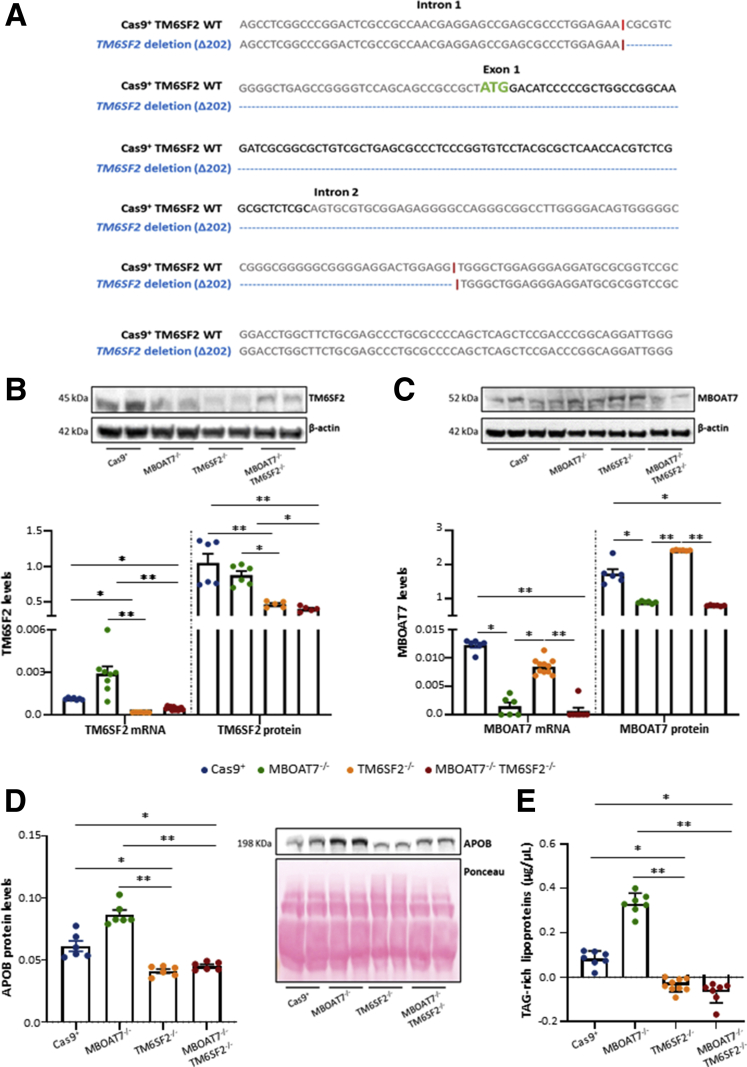
Methods: NAFLD patients (n = 1380), of whom 121 had HCC, were stratified with a semiquantitative score ranging from 0 to 3 according to the number of PNPLA3, TM6SF2, and MBOAT7 at-risk variants. TM6SF2 was silenced in HepG2 (TM6SF2-/-) and MBOAT7-/- (MBOAT7-/-TM6SF2-/-) through Clustered regularly interspaced short palindromic repeats and CRISPR-associated protein 9 (CRISPR/Cas9).
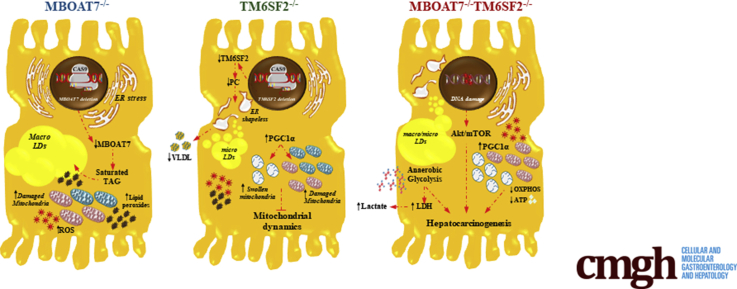
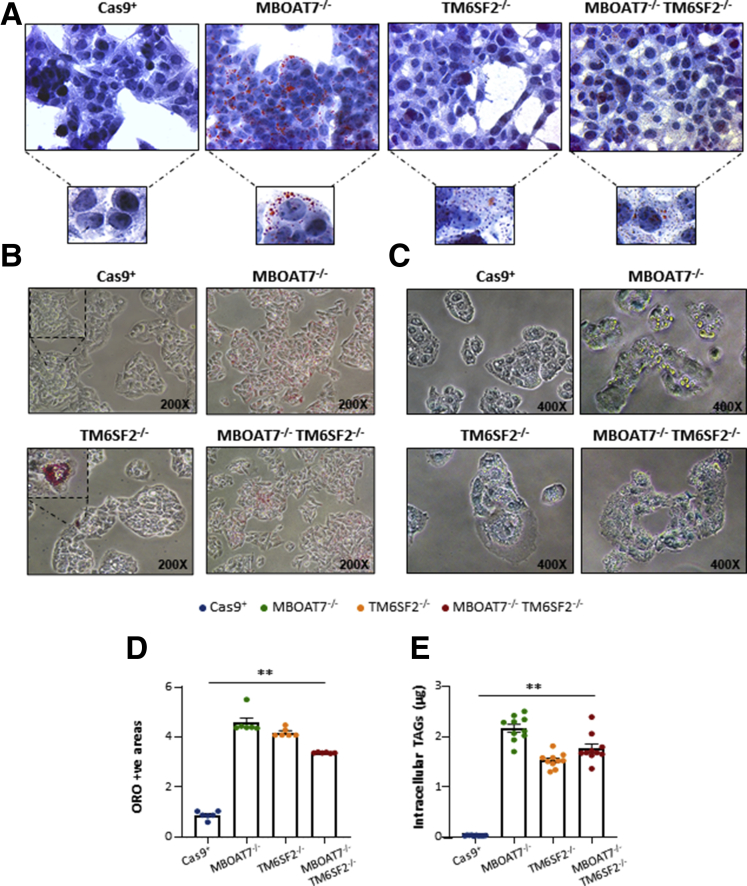
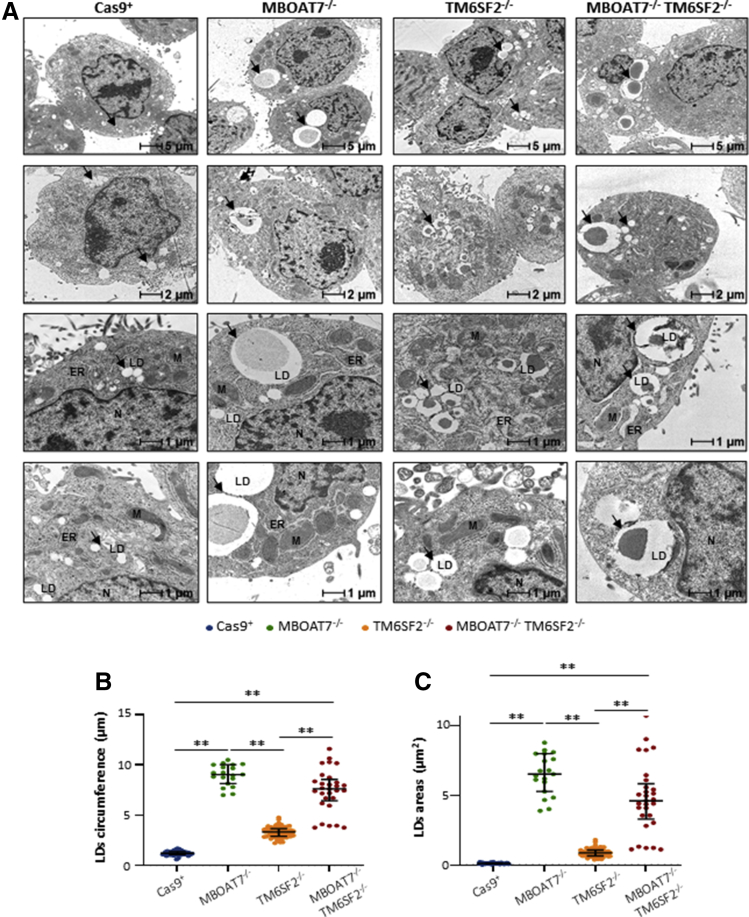
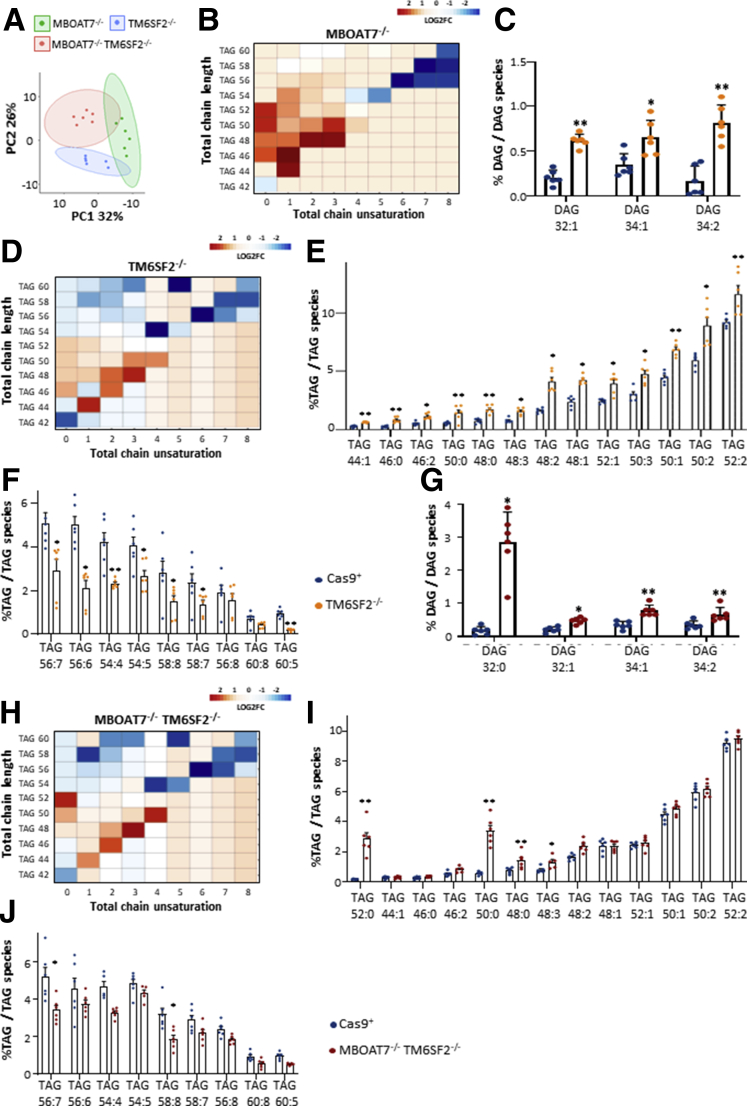
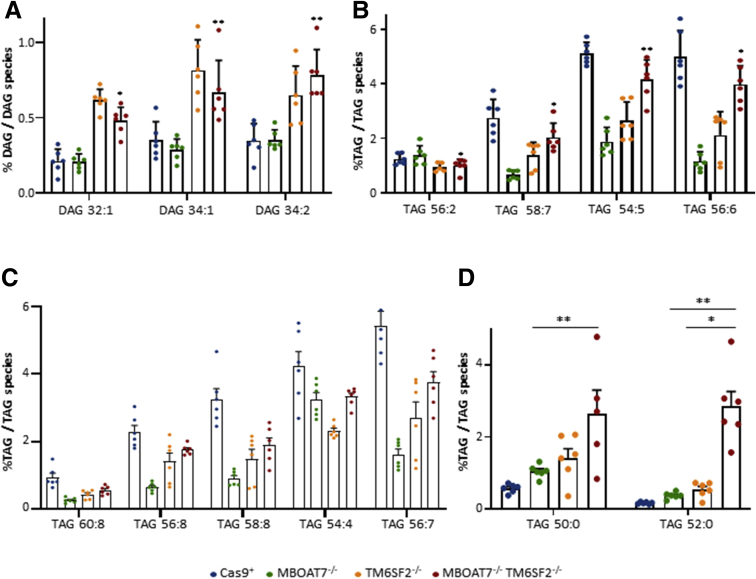
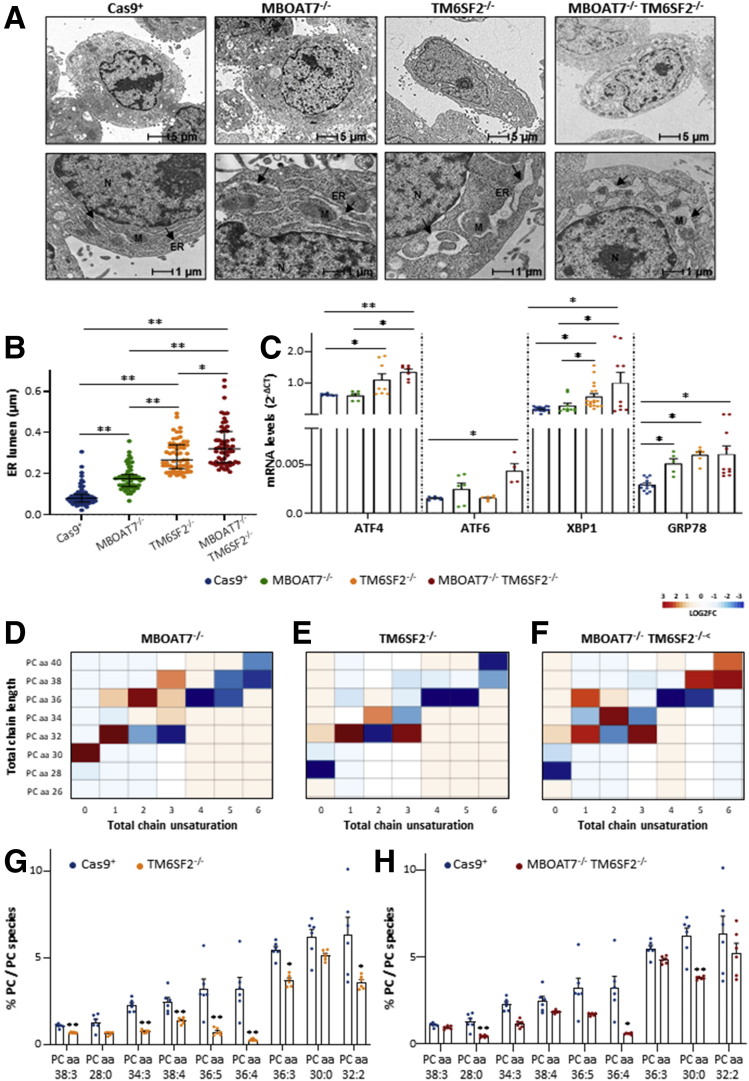
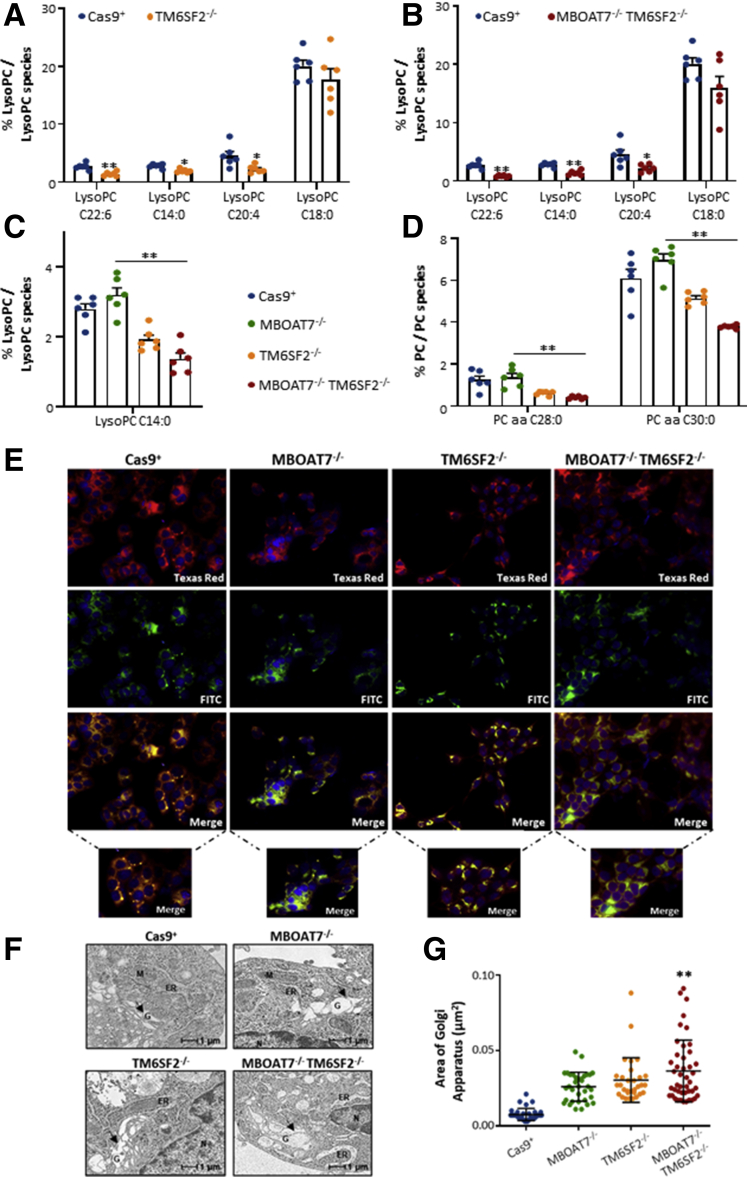
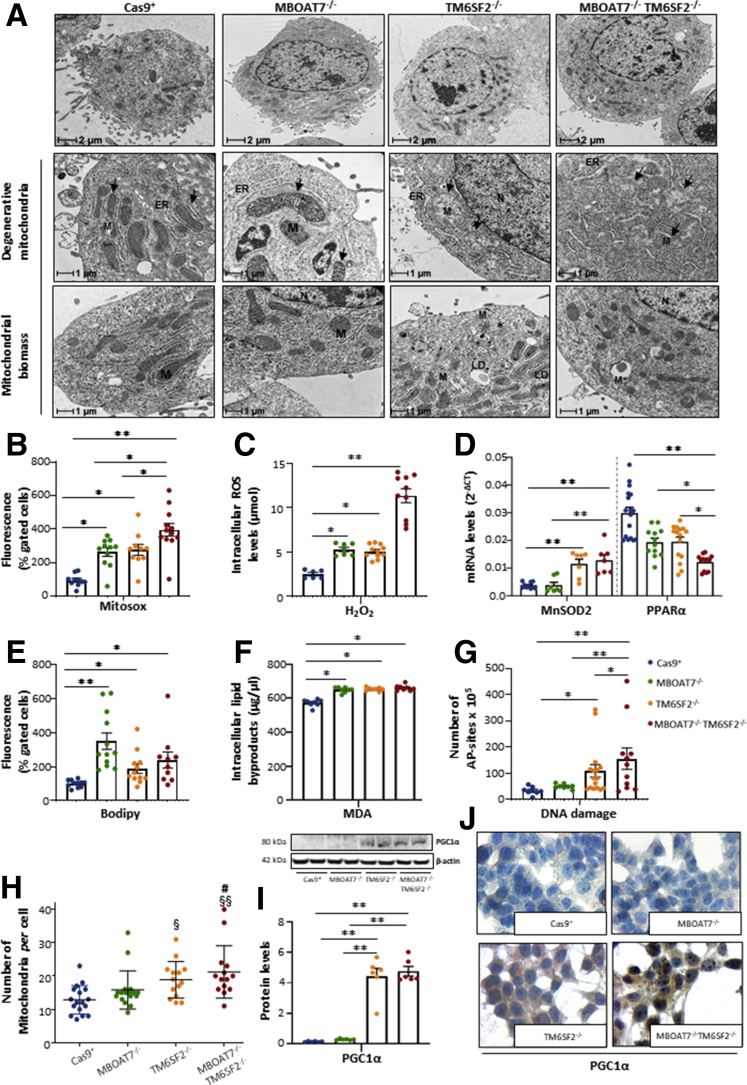
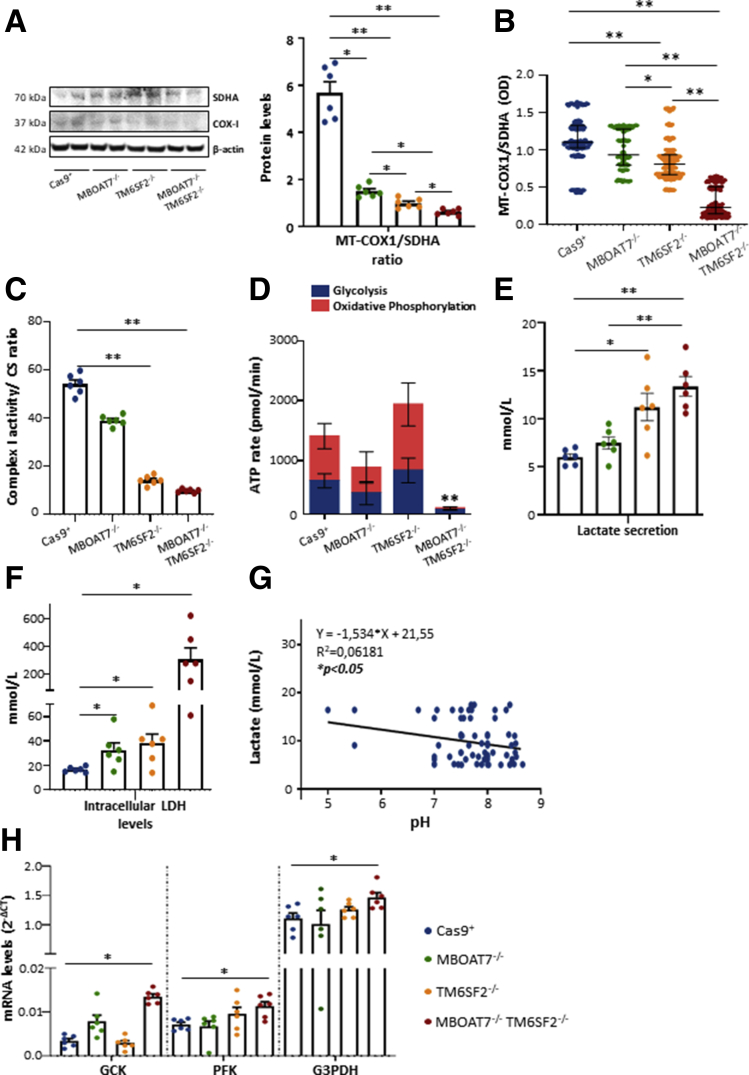
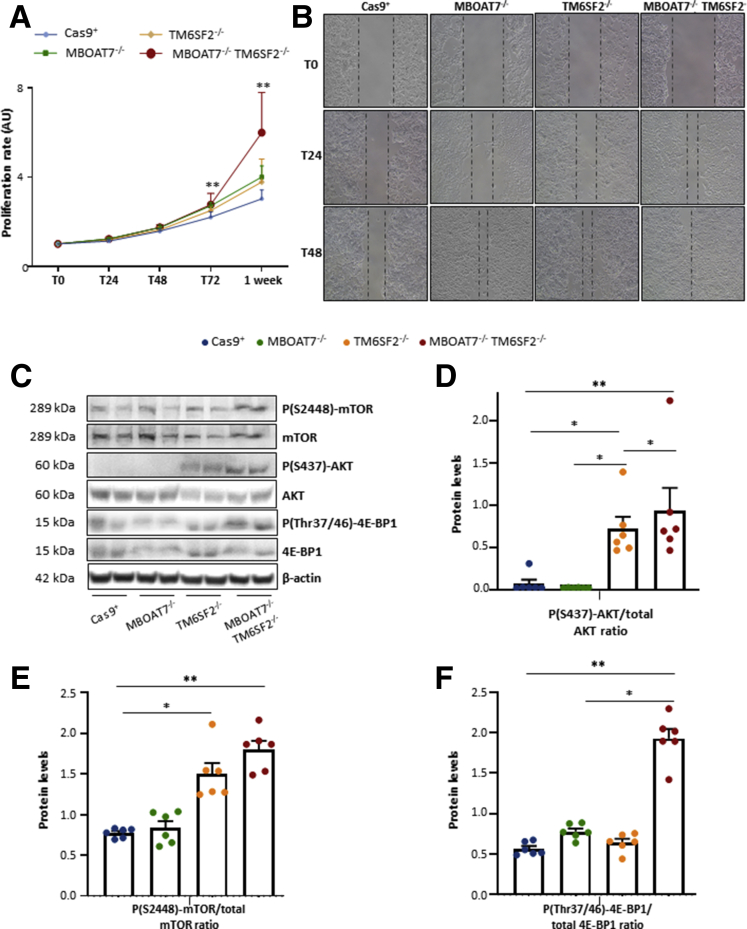
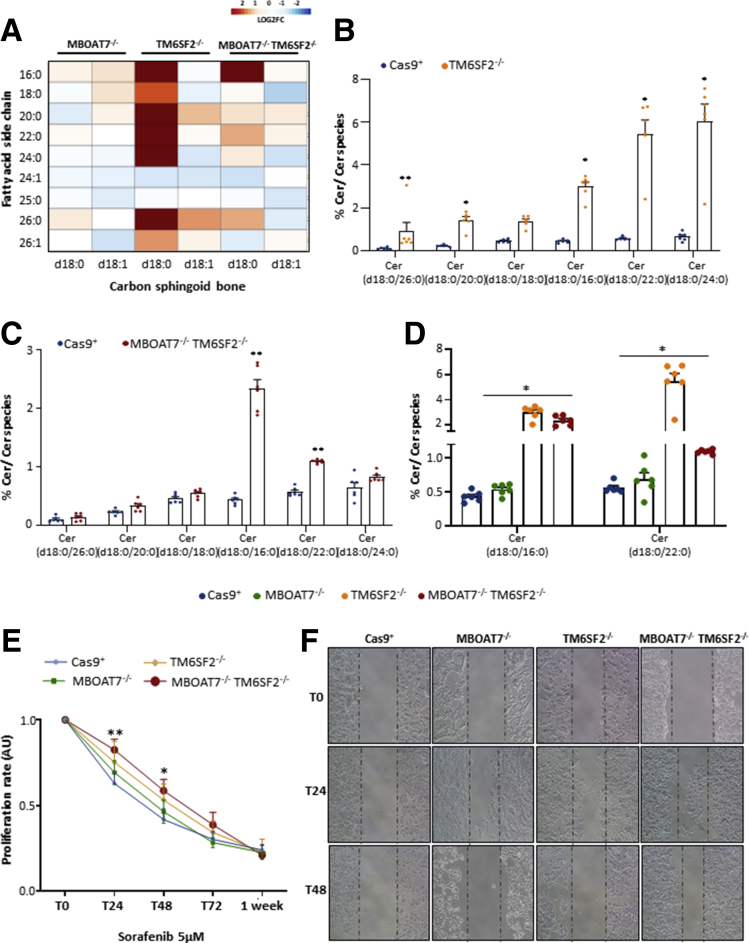
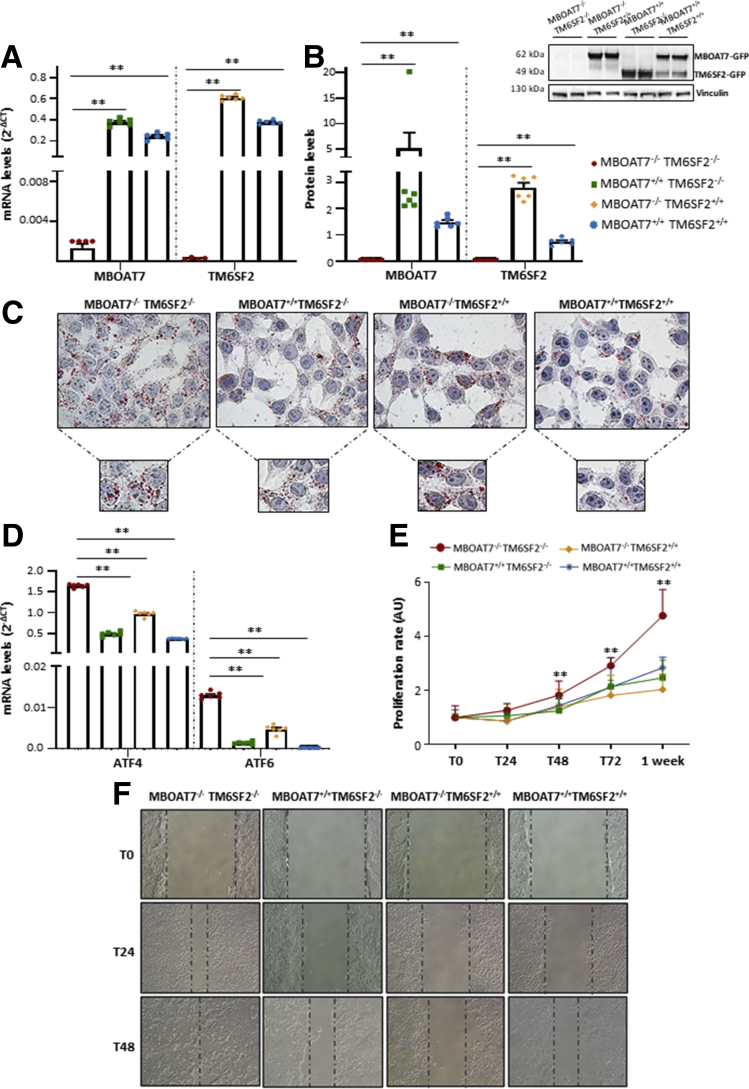
Results: In NAFLD patients, the additive weight of these mutations was associated with liver disease severity and an increased risk of developing HCC. In HepG2 cells, TM6SF2 silencing altered lipid composition and induced the accumulation of microvesicular lipid droplets (LDs), whereas the MBOAT7-/-TM6SF2-/- cells showed a mixed microvesicular/macrovesicular pattern of LDs. TM6SF2 deletion strongly affected endoplasmic reticulum and mitochondria ultrastructures, thus increasing endoplasmic reticulum/oxidative stress. The mitochondrial number was increased in both TM6SF2-/- and MBOAT7-/-TM6SF2-/- models, suggesting an unbalancing in mitochondrial dynamics, and the silencing of both MBOAT7 and TM6SF2 impaired mitochondrial activity with a shift toward anaerobic glycolysis. MBOAT7-/-TM6SF2-/- cells also showed the highest proliferation rate. Finally, the re-overexpression of MBOAT7 and/or TM6SF2 reversed the metabolic and tumorigenic features observed in the compound knockout model.
Conclusions: The co-presence of the 3 at-risk variants impacts the NAFLD course in both patients and experimental models, affecting LD accumulation, mitochondrial functionality, and metabolic reprogramming toward HCC.
Keywords: ER Stress; HCC; Mitochondrial Dynamics; NAFLD; TM6SF2.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
