

Plant LHC-like proteins show robust folding and static non-photochemical quenching
- PMID: 34824207
- PMCID: PMC8617258
- DOI: 10.1038/s41467-021-27155-1
Plant LHC-like proteins show robust folding and static non-photochemical quenching
Erratum in
-
Author Correction: Plant LHC-like proteins show robust folding and static non-photochemical quenching.Nat Commun. 2022 Mar 16;13(1):1528. doi: 10.1038/s41467-022-29099-6. Nat Commun. 2022. PMID: 35296680 Free PMC article. No abstract available.
Abstract
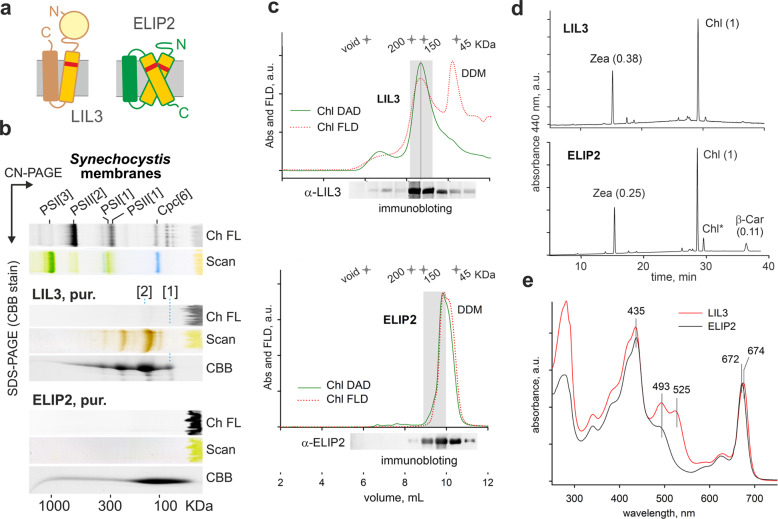
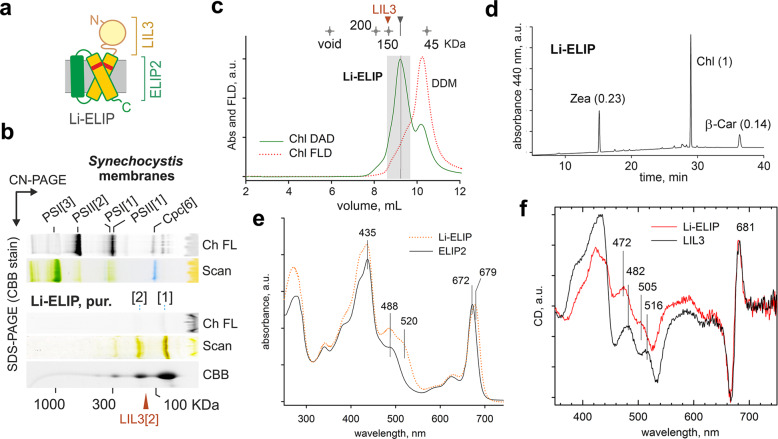
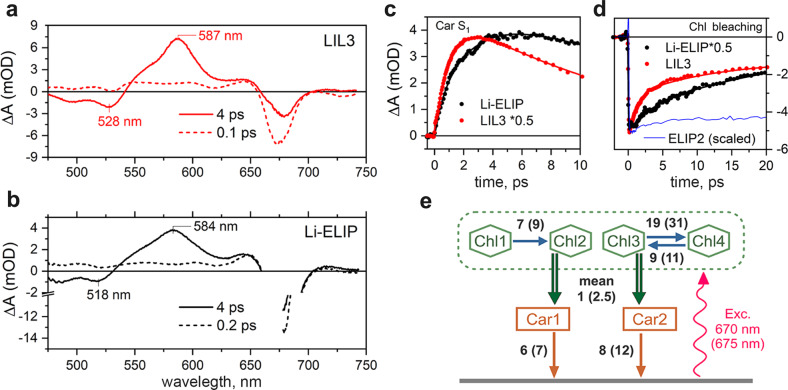
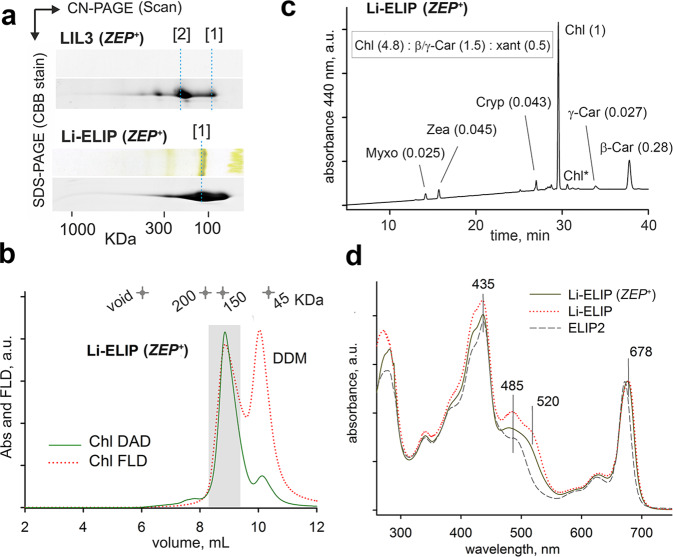
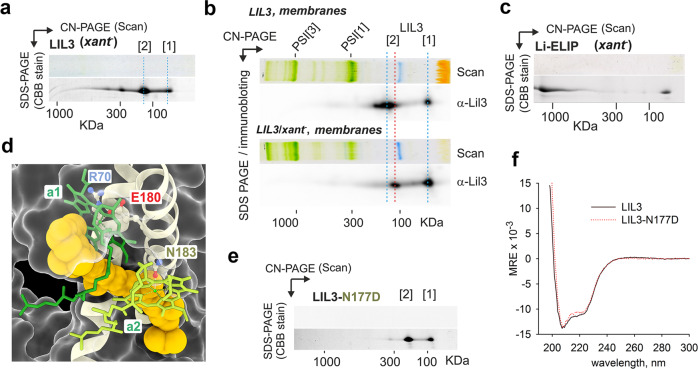
Life on Earth depends on photosynthesis, the conversion of light energy into chemical energy. Plants collect photons by light harvesting complexes (LHC)-abundant membrane proteins containing chlorophyll and xanthophyll molecules. LHC-like proteins are similar in their amino acid sequence to true LHC antennae, however, they rather serve a photoprotective function. Whether the LHC-like proteins bind pigments has remained unclear. Here, we characterize plant LHC-like proteins (LIL3 and ELIP2) produced in the cyanobacterium Synechocystis sp. PCC 6803 (hereafter Synechocystis). Both proteins were associated with chlorophyll a (Chl) and zeaxanthin and LIL3 was shown to be capable of quenching Chl fluorescence via direct energy transfer from the Chl Qy state to zeaxanthin S1 state. Interestingly, the ability of the ELIP2 protein to quench can be acquired by modifying its N-terminal sequence. By employing Synechocystis carotenoid mutants and site-directed mutagenesis we demonstrate that, although LIL3 does not need pigments for folding, pigments stabilize the LIL3 dimer.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Functional analysis of light-harvesting-like protein 3 (LIL3) and its light-harvesting chlorophyll-binding motif in Arabidopsis.J Biol Chem. 2014 Jan 10;289(2):987-99. doi: 10.1074/jbc.M113.525428. Epub 2013 Nov 25. J Biol Chem. 2014. PMID: 24275650 Free PMC article.
-
Xanthophyll carotenoids stabilise the association of cyanobacterial chlorophyll synthase with the LHC-like protein HliD.Biochem J. 2020 Oct 30;477(20):4021-4036. doi: 10.1042/BCJ20200561. Biochem J. 2020. PMID: 32990304
-
LIL3, a light-harvesting-like protein, plays an essential role in chlorophyll and tocopherol biosynthesis.Proc Natl Acad Sci U S A. 2010 Sep 21;107(38):16721-5. doi: 10.1073/pnas.1004699107. Epub 2010 Sep 7. Proc Natl Acad Sci U S A. 2010. PMID: 20823244 Free PMC article.
-
Energy transfer reactions involving carotenoids: quenching of chlorophyll fluorescence.J Photochem Photobiol B. 1996 Oct;36(1):3-15. doi: 10.1016/S1011-1344(96)07397-6. J Photochem Photobiol B. 1996. PMID: 8988608 Review.
-
Light harvesting complexes in chlorophyll c-containing algae.Biochim Biophys Acta Bioenerg. 2020 Apr 1;1861(4):148027. doi: 10.1016/j.bbabio.2019.05.003. Epub 2019 May 31. Biochim Biophys Acta Bioenerg. 2020. PMID: 31153887 Review.
Cited by
-
Moderate Temperature Reduction Changes the High-Light Acclimation Strategy of Lettuce Plants.Physiol Plant. 2025 May-Jun;177(3):e70298. doi: 10.1111/ppl.70298. Physiol Plant. 2025. PMID: 40457738 Free PMC article.
-
High-light-inducible proteins control associations between chlorophyll synthase and the Photosystem II biogenesis factor Ycf39.Plant Physiol. 2025 May 30;198(2):kiaf213. doi: 10.1093/plphys/kiaf213. Plant Physiol. 2025. PMID: 40413779 Free PMC article.
-
Protein Changes in Shade and Sun Haberlea rhodopensis Leaves during Dehydration at Optimal and Low Temperatures.Plants (Basel). 2023 Jan 15;12(2):401. doi: 10.3390/plants12020401. Plants (Basel). 2023. PMID: 36679114 Free PMC article.
-
High-resolution dissection of photosystem II electron transport reveals differential response to water deficit and heat stress in isolation and combination in pearl millet [Pennisetum glaucum (L.) R. Br.].Front Plant Sci. 2022 Aug 12;13:892676. doi: 10.3389/fpls.2022.892676. eCollection 2022. Front Plant Sci. 2022. PMID: 36035679 Free PMC article.
-
Coevolution of tandemly repeated hlips and RpaB-like transcriptional factor confers desiccation tolerance to subaerial Nostoc species.Proc Natl Acad Sci U S A. 2022 Oct 18;119(42):e2211244119. doi: 10.1073/pnas.2211244119. Epub 2022 Oct 10. Proc Natl Acad Sci U S A. 2022. PMID: 36215485 Free PMC article.
References
-
- Hashimoto H, Uragami C, Cogdell RJ. Carotenoids and photosynthesis. Subcell. Biochem. 2016;79:111–139. - PubMed
-
- Toporik H, Li J, Williams D, Chiu PL, Mazor Y. The structure of the stress-induced photosystem I-IsiA antenna supercomplex. Nat. Struct. Mol. Biol. 2019;26:443–449. - PubMed
-
- Shukla MK, et al. Binding of pigments to the cyanobacterial high-light-inducible protein HliC. Photosyn. Res. 2018;137:29–39. - PubMed
-
- Staleva H, et al. Mechanism of photoprotection in the cyanobacterial ancestor of plant antenna proteins. Nat. Chem. Biol. 2015;11:287–291. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
