

Andrographolide promotes hippocampal neurogenesis and spatial memory in the APPswe/PS1ΔE9 mouse model of Alzheimer's disease
- PMID: 34824314
- PMCID: PMC8616902
- DOI: 10.1038/s41598-021-01977-x
Andrographolide promotes hippocampal neurogenesis and spatial memory in the APPswe/PS1ΔE9 mouse model of Alzheimer's disease
Abstract
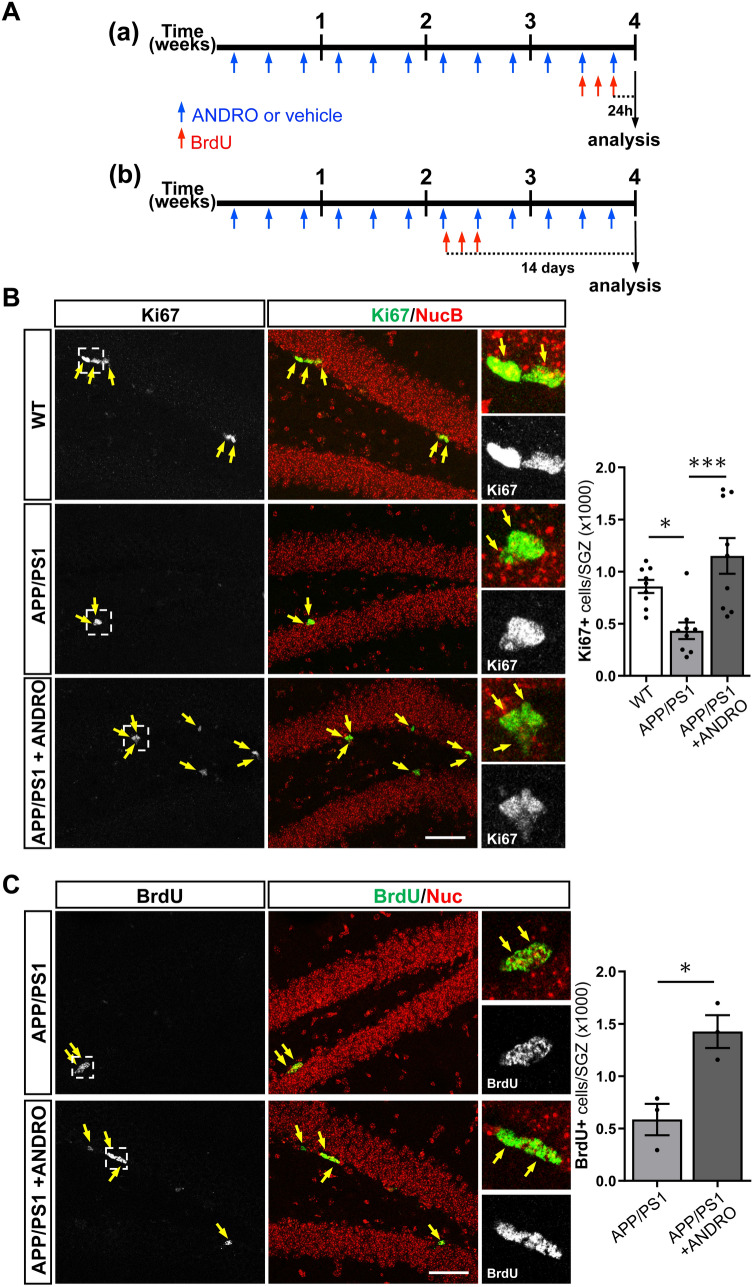
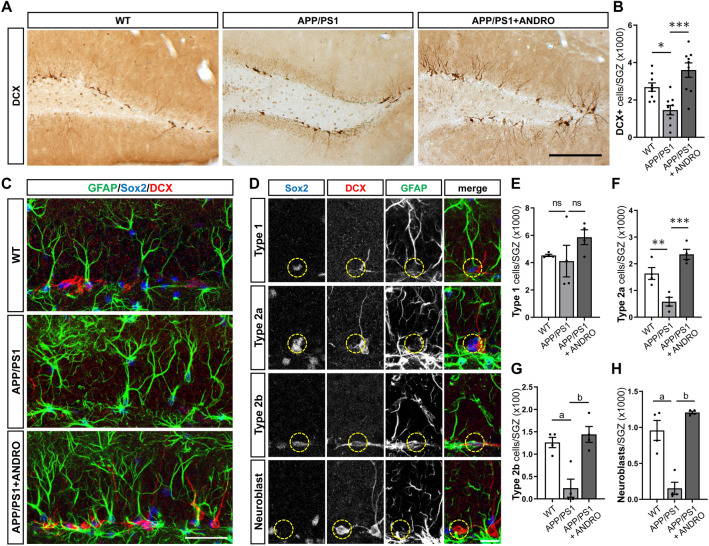
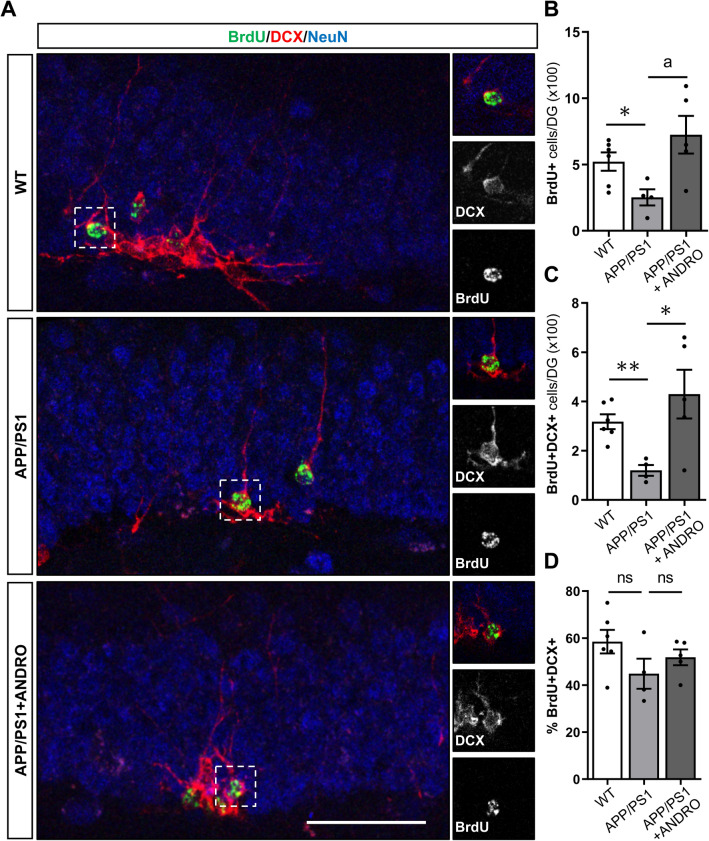
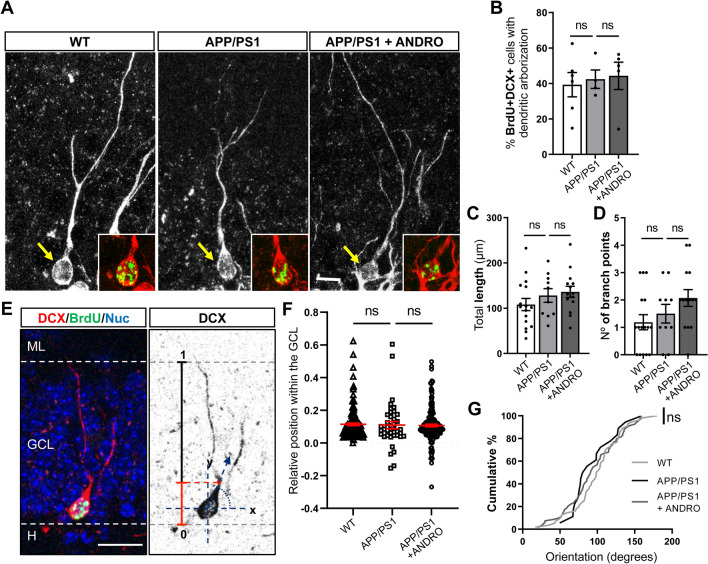
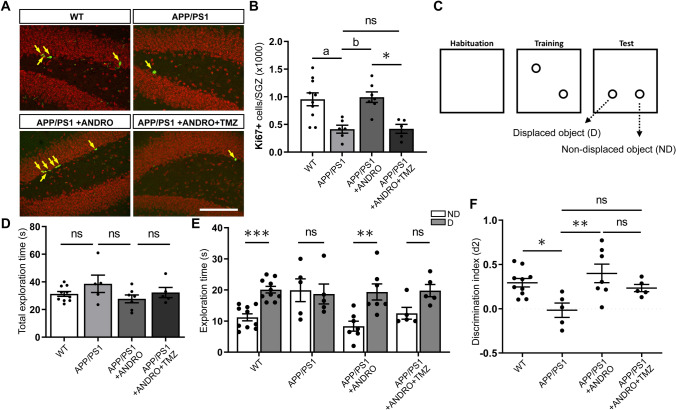
In Alzheimer´s disease (AD) there is a reduction in hippocampal neurogenesis that has been associated to cognitive deficits. Previously we showed that Andrographolide (ANDRO), the main bioactive component of Andrographis paniculate, induces proliferation in the hippocampus of the APPswe/PSEN1ΔE9 (APP/PS1) mouse model of AD as assessed by staining with the mitotic marker Ki67. Here, we further characterized the effect of ANDRO on hippocampal neurogenesis in APP/PS1 mice and evaluated the contribution of this process to the cognitive effect of ANDRO. Treatment of 8-month-old APP/PS1 mice with ANDRO for 4 weeks increased proliferation in the dentate gyrus as evaluated by BrdU incorporation. Although ANDRO had no effect on neuronal differentiation of newborn cells, it strongly increased neural progenitors, neuroblasts and newborn immature neurons, cell populations that were decreased in APP/PS1 mice compared to age-matched wild-type mice. ANDRO had no effect on migration or in total dendritic length, arborization and orientation of immature neurons, suggesting no effects on early morphological development of newborn neurons. Finally, ANDRO treatment improved the performance of APP/PS1 mice in the object location memory task. This effect was not completely prevented by co-treatment with the anti-mitotic drug TMZ, suggesting that other effects of ANDRO in addition to the increase in neurogenesis might underlie the observed cognitive improvement. Altogether, our data indicate that in APP/PS1 mice ANDRO stimulates neurogenesis in the hippocampus by inducing proliferation of neural precursor cells and improves spatial memory performance.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
