

Germline biallelic mutation affecting the transcription factor Helios causes pleiotropic defects of immunity
- PMID: 34826259
- PMCID: PMC7612971
- DOI: 10.1126/sciimmunol.abe3981
Germline biallelic mutation affecting the transcription factor Helios causes pleiotropic defects of immunity
Abstract
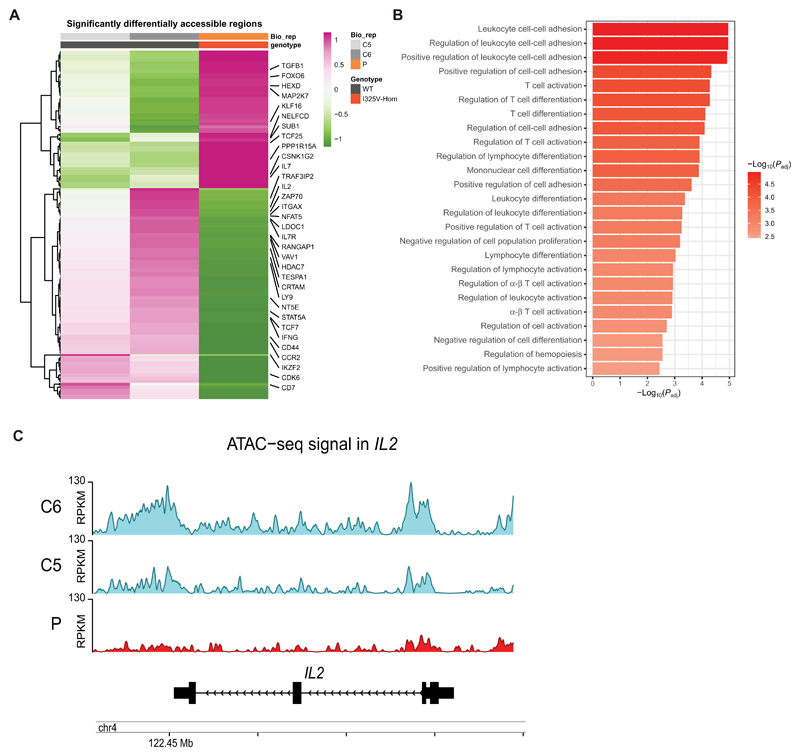
Helios, a member of the Ikaros family of transcription factors, is predominantly expressed in developing thymocytes, activated T cells, and regulatory T cells (Tregs). Studies in mice have emphasized its role in maintenance of Treg immunosuppressive functions by stabilizing Foxp3 expression and silencing the Il2 locus. However, its contribution to human immune homeostasis and the precise mechanisms by which Helios regulates other T cell subsets remain unresolved. Here, we investigated a patient with recurrent respiratory infections and hypogammaglobulinemia and identified a germline homozygous missense mutation in IKZF2 encoding Helios (p.Ile325Val). We found that HeliosI325V retains DNA binding and dimerization properties but loses interaction with several partners, including epigenetic remodelers. Whereas patient Tregs showed increased IL-2 production, patient conventional T cells had decreased accessibility of the IL2 locus and consequently reduced IL-2 production. Reduced chromatin accessibility was not exclusive to the IL2 locus but involved a variety of genes associated with T cell activation. Single-cell RNA sequencing of peripheral blood mononuclear cells revealed gene expression signatures indicative of a shift toward a proinflammatory, effector-like status in patient CD8+ T cells. Moreover, patient CD4+ T cells exhibited a pronounced defect in proliferation with delayed expression of surface checkpoint inhibitors, suggesting an impaired onset of the T cell activation program. Collectively, we identified a previously uncharacterized, germline-encoded inborn error of immunity and uncovered a cell-specific defect in Helios-dependent epigenetic regulation. Binding of Helios with specific partners mediates this regulation, which is ultimately necessary for the transcriptional programs that enable T cell homeostasis in health and disease.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures






References
-
- John LB, Ward AC. The Ikaros gene family: Transcriptional regulators of hematopoiesis and immunity. Mol Immunol. 2011;48:1272–1278. - PubMed
-
- Heizmann B, Kastner P, Chan S. The Ikaros family in lymphocyte development. Curr Opin Immunol. 2018;51:14–23. - PubMed
-
- Sridharan R, Smale ST. Predominant interaction of both Ikaros and Helios with the NuRD complex in immature thymocytes. J Biol Chem. 2007;282:30227–30238. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
