

Carrot AOX2a Transcript Profile Responds to Growth and Chilling Exposure
- PMID: 34834732
- PMCID: PMC8625938
- DOI: 10.3390/plants10112369
Carrot AOX2a Transcript Profile Responds to Growth and Chilling Exposure
Abstract
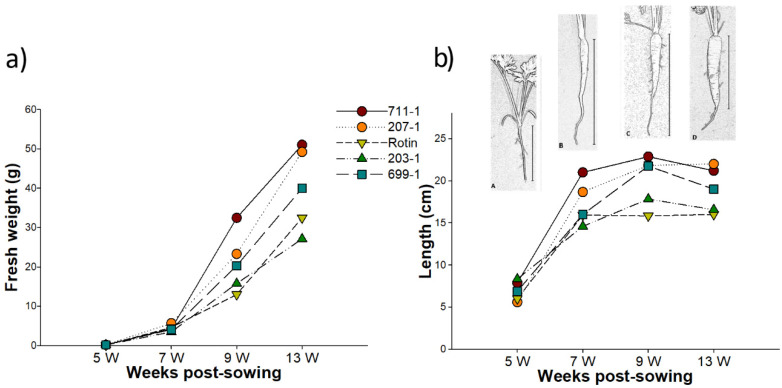
Alternative oxidase (AOX) is a key enzyme of the alternative respiration, known to be involved in plant development and in response to various stresses. To verify the role of DcAOX1 and DcAOX2a genes in carrot tap root growth and in response to cold stress, their expression was analyzed in two experiments: during root growth for 13 weeks and in response to a cold challenge trial of 7 days, in both cases using different carrot cultivars. Carrot root growth is initially characterized by an increase in length, followed by a strong increase in weight. DcAOX2a presented the highest expression levels during the initial stages of root growth for all cultivars, but DcAOX1 showed no particular trend in expression. Cold stress had a negative impact on root growth, and generally up-regulated DcAOX2a with no consistent effect on DcAOX1. The identification of cis-acting regulatory elements (CAREs) located at the promoters of both genes showed putative sequences involved in cold stress responsiveness, as well as growth. However, DcAOX2a promoter presented more CAREs related to hormonal pathways, including abscisic acid and gibberellins synthesis, than DcAOX1. These results point to a dual role of DcAOX2a on carrot tap root secondary growth and cold stress response.
Keywords: Daucus carota; alternative oxidase; chilling stress; cis-elements; growth.
Conflict of interest statement
The authors declare no conflict of interest.
Figures








References
-
- Finnegan P.M., Soole K.L., Umbach A.L. Advances in Photosynthesis and Respiration. Plant Mitochondria: From Genome to Function. Springer; Dordrecht, The Netherlands: 2004. Alternative mitochondrial electron transport proteins in higher plants; pp. 163–230.
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
