

Nucleation, stabilization, and disassembly of branched actin networks
- PMID: 34836783
- PMCID: PMC9018471
- DOI: 10.1016/j.tcb.2021.10.006
Nucleation, stabilization, and disassembly of branched actin networks
Abstract
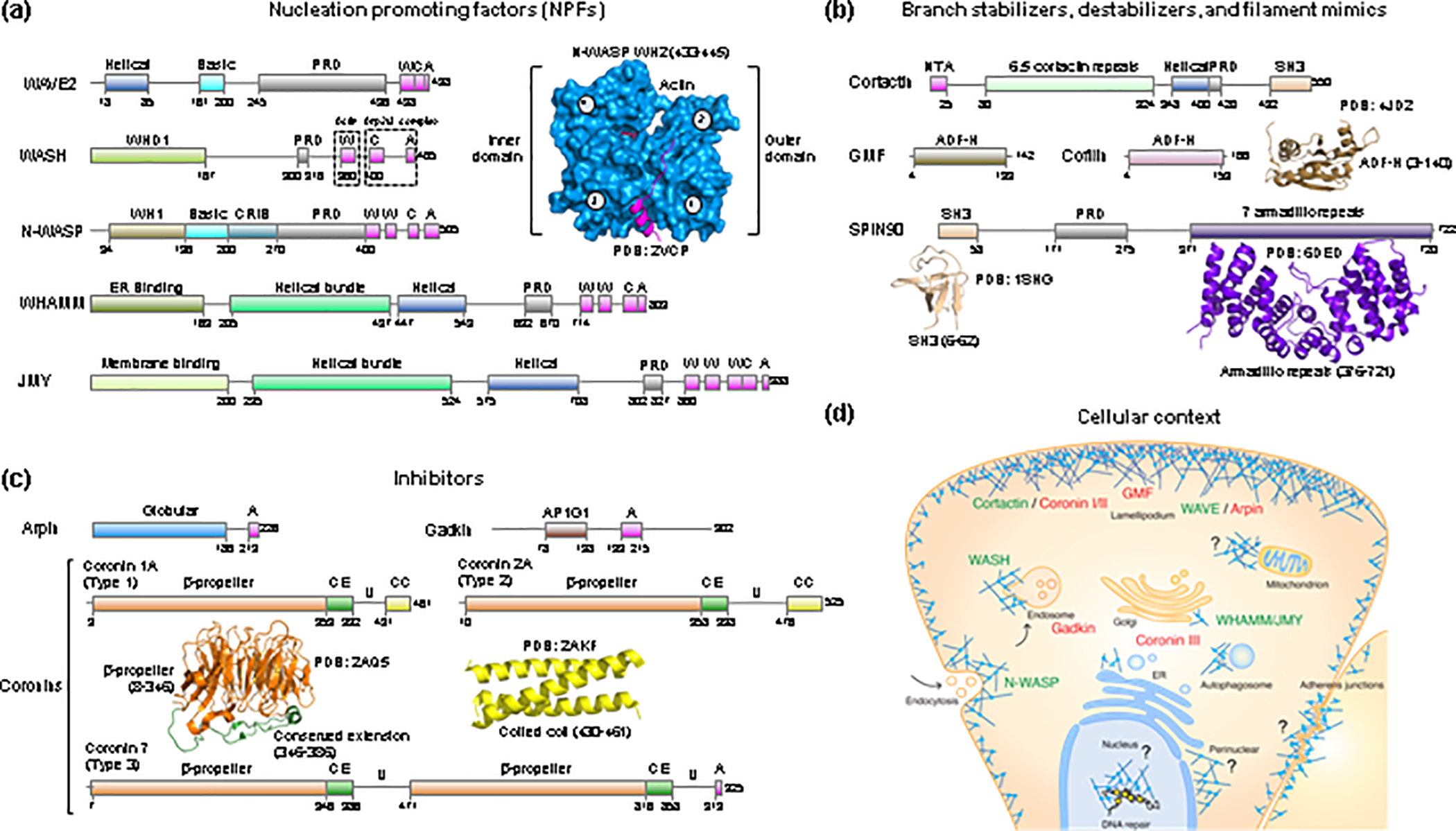
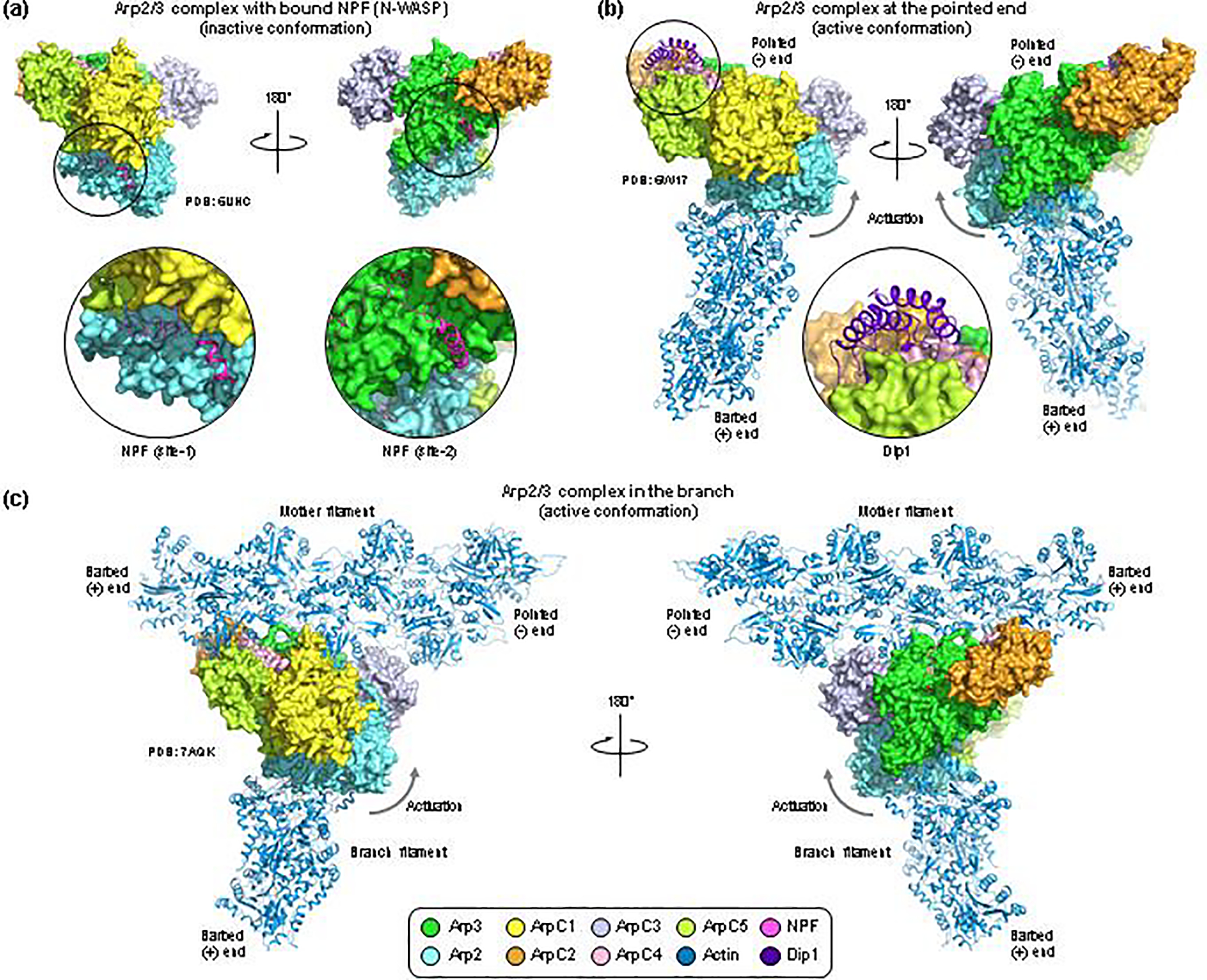
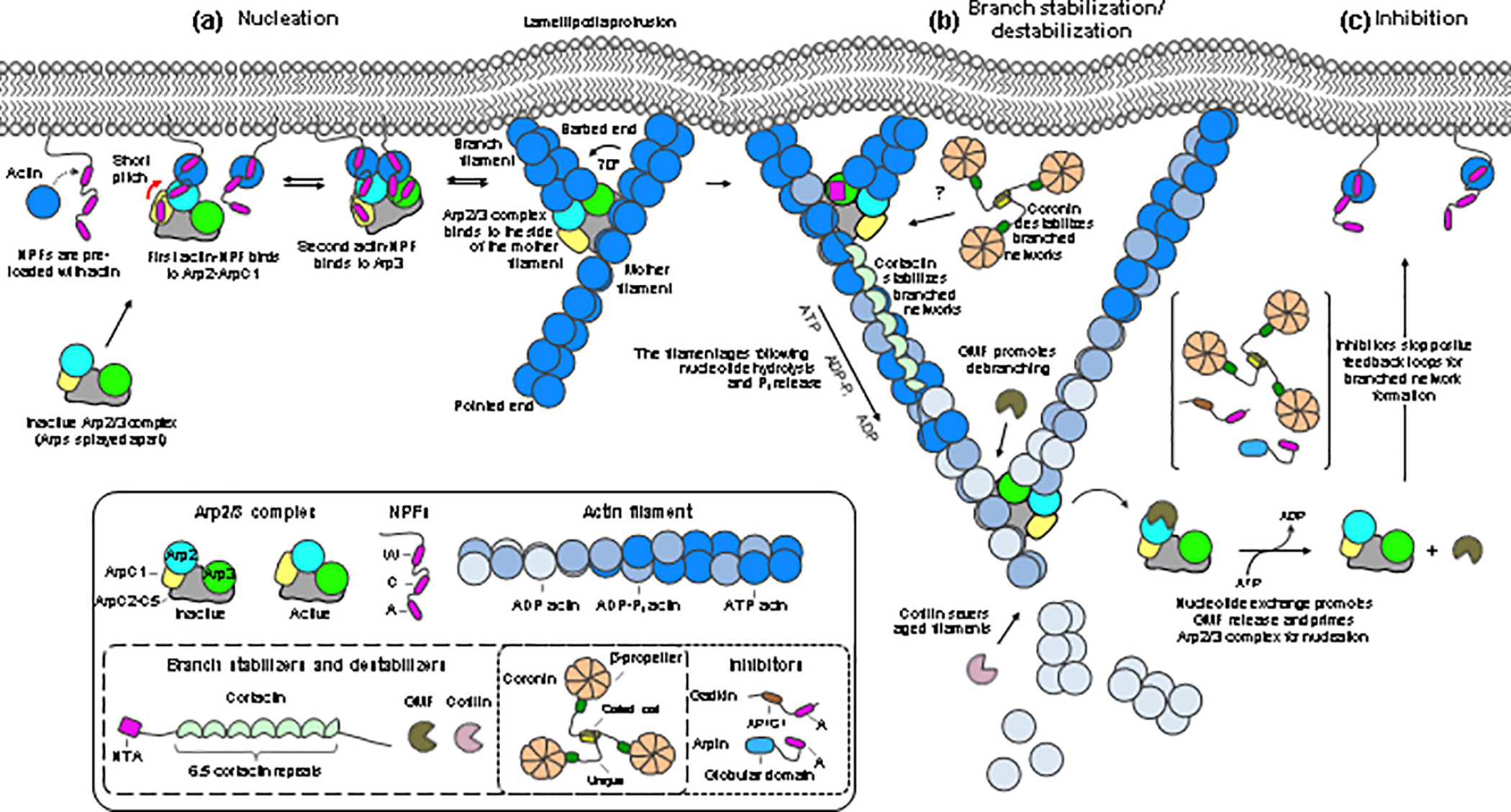
Arp2/3 complex is an actin filament nucleation and branching machinery conserved in all eukaryotes from yeast to human. Arp2/3 complex branched networks generate pushing forces that drive cellular processes ranging from membrane remodeling to cell and organelle motility. Several molecules regulate these processes by directly inhibiting or activating Arp2/3 complex and by stabilizing or disassembling branched networks. Here, we review recent advances in our understanding of Arp2/3 complex regulation, including high-resolution cryoelectron microscopy (cryo-EM) structures that illuminate the mechanisms of Arp2/3 complex activation and branch formation, and novel cellular pathways of branch formation, stabilization, and debranching. We also identify major gaps in our understanding of Arp2/3 complex inhibition and branch stabilization and disassembly.
Keywords: Arp2/3 complex; branched network stabilization and disassembly; inhibitors; mechanosensation; nucleation-promoting factors.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Robinson RC et al. (2001) Crystal structure of Arp2/3 complex. Science 294 (5547), 1679–84. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
