

Infection and transmission of ancestral SARS-CoV-2 and its alpha variant in pregnant white-tailed deer
- PMID: 34842046
- PMCID: PMC8725908
- DOI: 10.1080/22221751.2021.2012528
Infection and transmission of ancestral SARS-CoV-2 and its alpha variant in pregnant white-tailed deer
Abstract
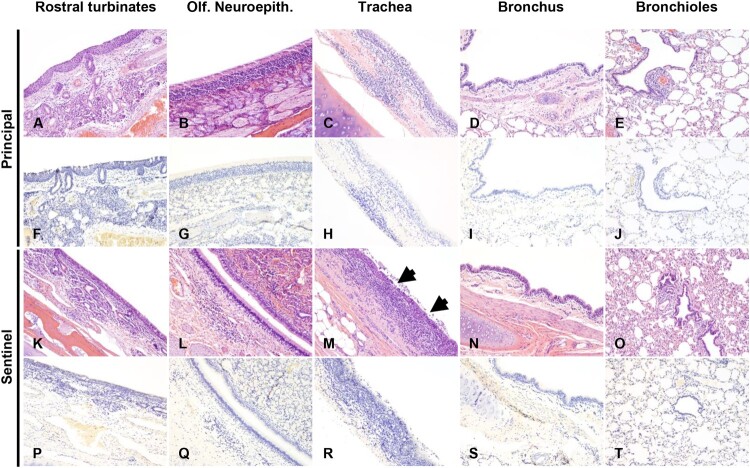
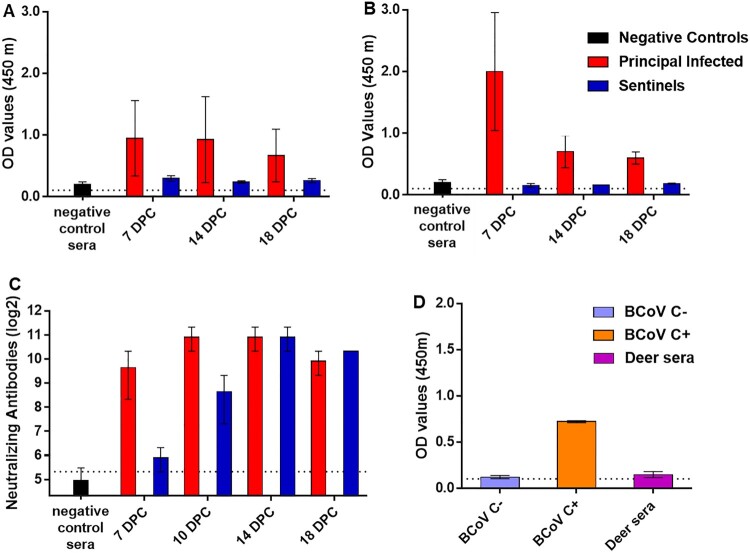
ABSTRACTSARS-CoV-2 was first reported circulating in human populations in December 2019 and has since become a global pandemic. Recent history involving SARS-like coronavirus outbreaks have demonstrated the significant role of intermediate hosts in viral maintenance and transmission. Evidence of SARS-CoV-2 natural infection and experimental infections of a wide variety of animal species has been demonstrated, and in silico and in vitro studies have indicated that deer are susceptible to SARS-CoV-2 infection. White-tailed deer (WTD) are amongst the most abundant and geographically widespread wild ruminant species in the US. Recently, WTD fawns were shown to be susceptible to SARS-CoV-2. In the present study, we investigated the susceptibility and transmission of SARS-CoV-2 in adult WTD. In addition, we examined the competition of two SARS-CoV-2 isolates, representatives of the ancestral lineage A and the alpha variant of concern (VOC) B.1.1.7 through co-infection of WTD. Next-generation sequencing was used to determine the presence and transmission of each strain in the co-infected and contact sentinel animals. Our results demonstrate that adult WTD are highly susceptible to SARS-CoV-2 infection and can transmit the virus through direct contact as well as vertically from doe to fetus. Additionally, we determined that the alpha VOC B.1.1.7 isolate of SARS-CoV-2 outcompetes the ancestral lineage A isolate in WTD, as demonstrated by the genome of the virus shed from nasal and oral cavities from principal infected and contact animals, and from the genome of virus present in tissues of principal infected deer, fetuses and contact animals.
Keywords: SARS-CoV-2; cervid; co-infection; pregnancy; susceptibility; transmission; white-tailed deer.
Conflict of interest statement
Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture. USDA is an equal opportunity provider and employer. The J.A.R laboratory received support from Tonix Pharmaceuticals, Xing Technologies and Zoetis, outside of the reported work. J.A.R. is inventor on patents and patent applications on the use of antivirals and vaccines for the treatment and prevention of virus infections, owned by Kansas State University, KS, or the Icahn School of Medicine at Mount Sinai, New York. The A.G.-S. laboratory has received research support from Pfizer, Senhwa Biosciences, Kenall Manufacturing, Avimex, Johnson & Johnson, Dynavax, 7Hills Pharma, Pharmamar, ImmunityBio, Accurius, Nanocomposix, Hexamer, N-fold LLC, and Merck, outside of the reported work. A.G.-S. has consulting agreements for the following companies involving cash and/or stock: Vivaldi Biosciences, Contrafect, 7Hills Pharma, Avimex, Vaxalto, Pagoda, Accurius, Esperovax, Farmak, Applied Biological Laboratories and Pfizer, outside of the reported work. A.G.-S. is inventor on patents and patent applications on the use of antivirals and vaccines for the treatment and prevention of virus infections, owned by the Icahn School of Medicine at Mount Sinai, New York.
Figures
Update of
-
Infection and transmission of ancestral SARS-CoV-2 and its alpha variant in pregnant white-tailed deer.bioRxiv [Preprint]. 2021 Aug 24:2021.08.15.456341. doi: 10.1101/2021.08.15.456341. bioRxiv. 2021. Update in: Emerg Microbes Infect. 2022 Dec;11(1):95-112. doi: 10.1080/22221751.2021.2012528. PMID: 34426811 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous