

Redox conditions correlated with vibronic coupling modulate quantum beats in photosynthetic pigment-protein complexes
- PMID: 34845027
- PMCID: PMC8670468
- DOI: 10.1073/pnas.2112817118
Redox conditions correlated with vibronic coupling modulate quantum beats in photosynthetic pigment-protein complexes
Abstract
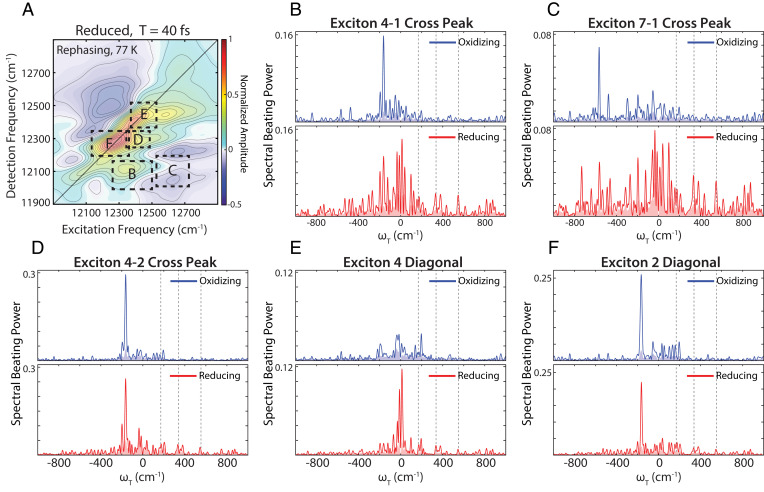
Quantum coherences, observed as time-dependent beats in ultrafast spectroscopic experiments, arise when light-matter interactions prepare systems in superpositions of states with differing energy and fixed phase across the ensemble. Such coherences have been observed in photosynthetic systems following ultrafast laser excitation, but what these coherences imply about the underlying energy transfer dynamics remains subject to debate. Recent work showed that redox conditions tune vibronic coupling in the Fenna-Matthews-Olson (FMO) pigment-protein complex in green sulfur bacteria, raising the question of whether redox conditions may also affect the long-lived (>100 fs) quantum coherences observed in this complex. In this work, we perform ultrafast two-dimensional electronic spectroscopy measurements on the FMO complex under both oxidizing and reducing conditions. We observe that many excited-state coherences are exclusively present in reducing conditions and are absent or attenuated in oxidizing conditions. Reducing conditions mimic the natural conditions of the complex more closely. Further, the presence of these coherences correlates with the vibronic coupling that produces faster, more efficient energy transfer through the complex under reducing conditions. The growth of coherences across the waiting time and the number of beating frequencies across hundreds of wavenumbers in the power spectra suggest that the beats are excited-state coherences with a mostly vibrational character whose phase relationship is maintained through the energy transfer process. Our results suggest that excitonic energy transfer proceeds through a coherent mechanism in this complex and that the coherences may provide a tool to disentangle coherent relaxation from energy transfer driven by stochastic environmental fluctuations.
Keywords: excitonics; light harvesting; photosynthesis; quantum biology; ultrafast spectroscopy.
Conflict of interest statement
The authors declare no competing interest.
Figures



References
-
- Blankenship R. E., Molecular Mechanisms of Photosynthesis (Wiley/Blackwell, Chichester, UK, ed. 2, 2014).
-
- Ruban A. V., Plant science: Crops on the fast track for light. Nature 541, 36–37 (2017). - PubMed
-
- Liu Z., et al. , Crystal structure of spinach major light-harvesting complex at 2.72 A resolution. Nature 428, 287–292 (2004). - PubMed
-
- Fenna R., Matthews B., Chlorophyll arrangement in a bacteriochlorophyll protein from Chlorobium limicola. Nature 258, 573–577 (1975).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
