

Branched-chain amino acids govern the high learning ability phenotype in Tokai high avoider (THA) rats
- PMID: 34845278
- PMCID: PMC8630195
- DOI: 10.1038/s41598-021-02591-7
Branched-chain amino acids govern the high learning ability phenotype in Tokai high avoider (THA) rats
Abstract
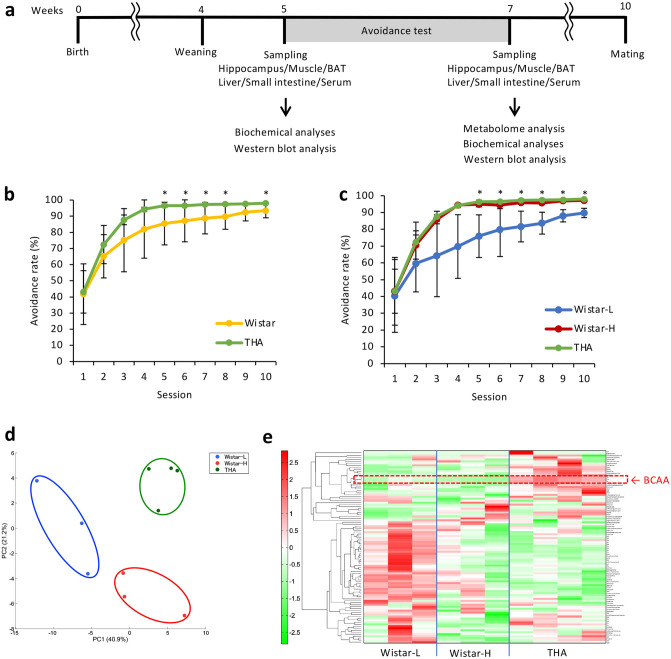
To fully understand the mechanisms governing learning and memory, animal models with minor interindividual variability and higher cognitive function are required. THA rats established by crossing those with high learning capacity exhibit excellent learning and memory abilities, but the factors underlying their phenotype are completely unknown. In the current study, we compare the hippocampi of parental strain Wistar rats to those of THA rats via metabolomic analysis in order to identify molecules specific to the THA rat hippocampus. Higher branched-chain amino acid (BCAA) levels and enhanced activation of BCAA metabolism-associated enzymes were observed in THA rats, suggesting that acetyl-CoA and acetylcholine are synthesized through BCAA catabolism. THA rats maintained high blood BCAA levels via uptake of BCAAs in the small intestine and suppression of BCAA catabolism in the liver. Feeding THA rats with a BCAA-reduced diet decreased acetylcholine levels and learning ability, thus, maintaining high BCAA levels while their proper metabolism in the hippocampus is the mechanisms underlying the high learning ability in THA rats. Identifying appropriate BCAA nutritional supplements and activation methods may thus hold potential for the prevention and amelioration of higher brain dysfunction, including learning disabilities and dementia.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Shigeta S, et al. Strain difference in learning impairment in rats following lead administration during brain development. Tokai. J. Exp. Clin. Med. 1986;11:241–247. - PubMed
-
- Shigeta S, et al. Neurobehavioral analysis of high-rate Sidman avoidance rat strain. Yakubutsu Seishin Kodo. 1989;9:217–224. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
