

Optical tweezers in single-molecule biophysics
- PMID: 34849486
- PMCID: PMC8629167
- DOI: 10.1038/s43586-021-00021-6
Optical tweezers in single-molecule biophysics
Abstract
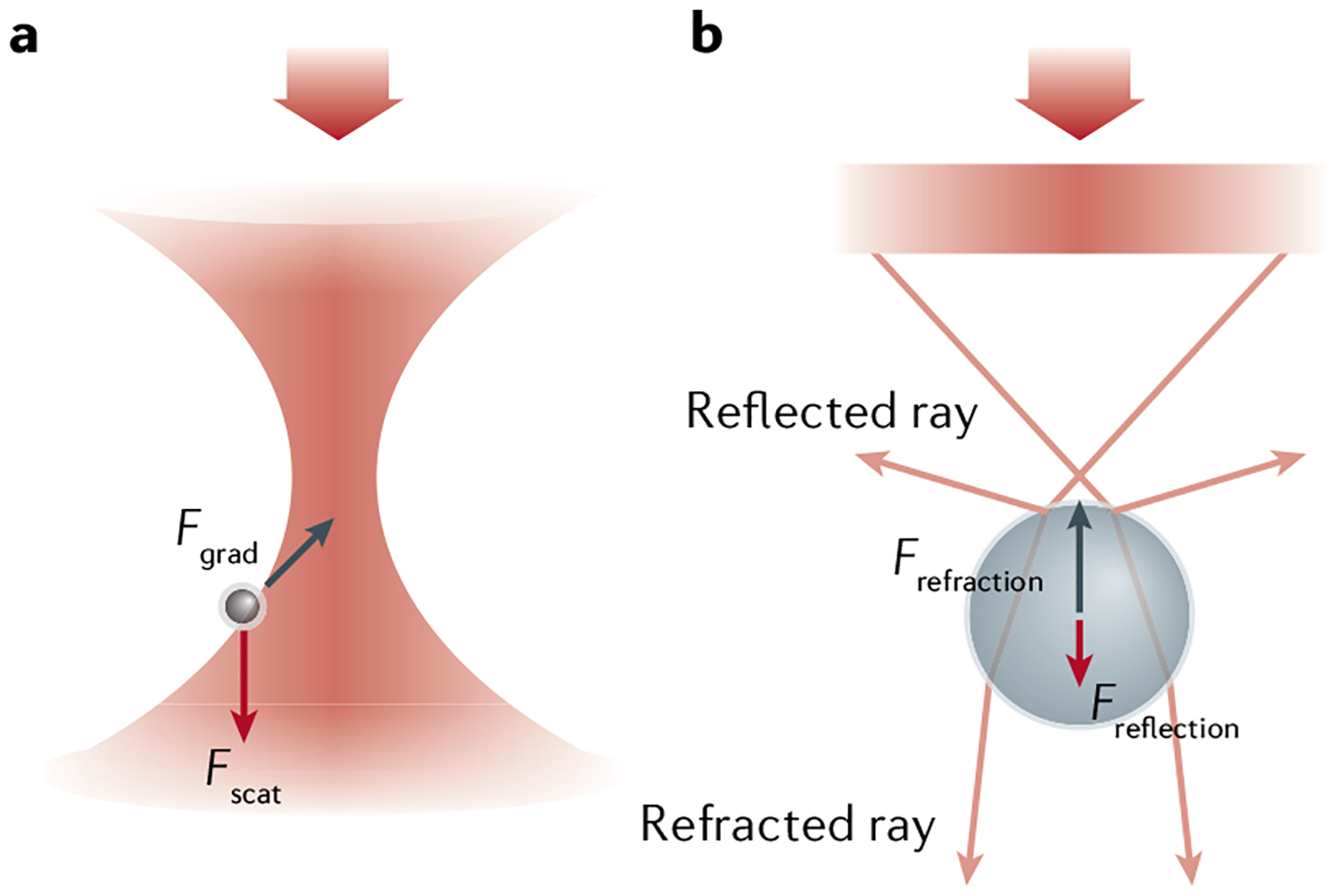
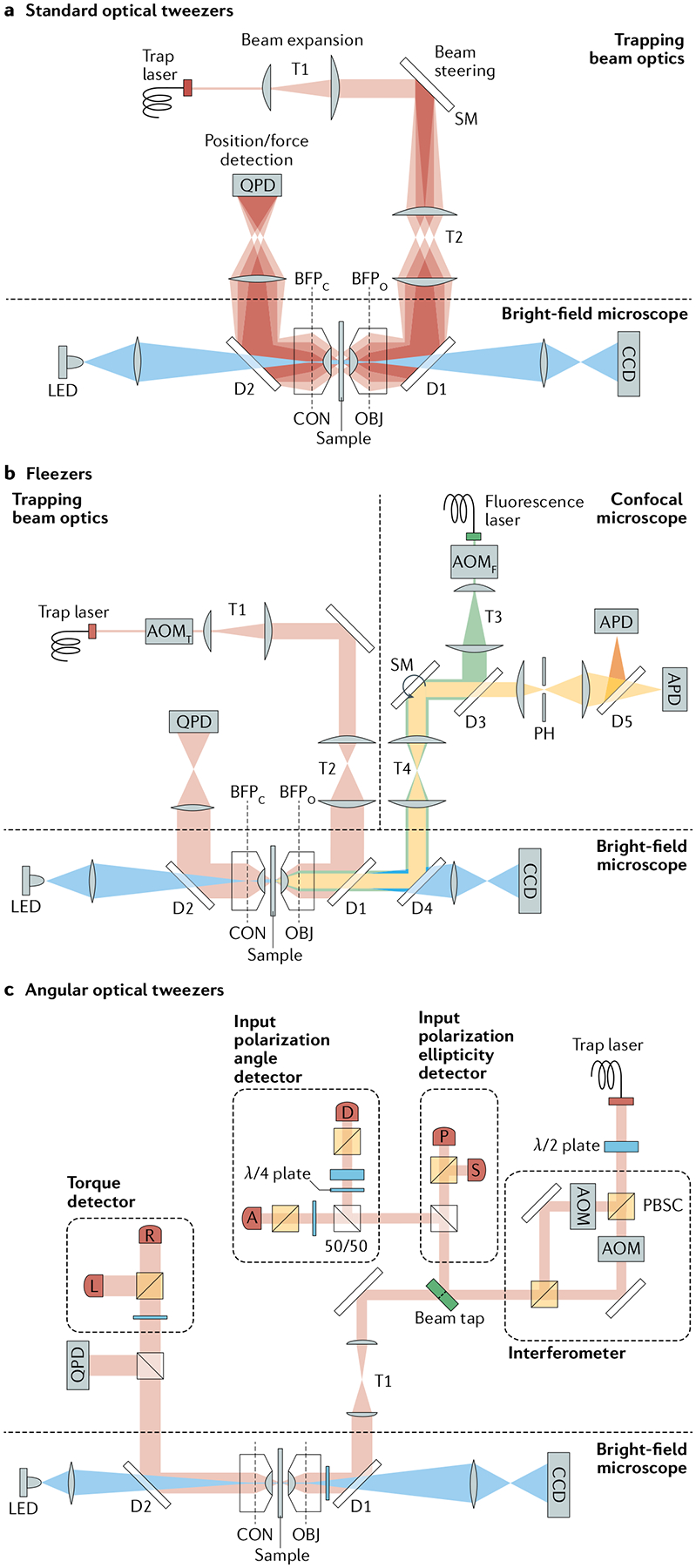
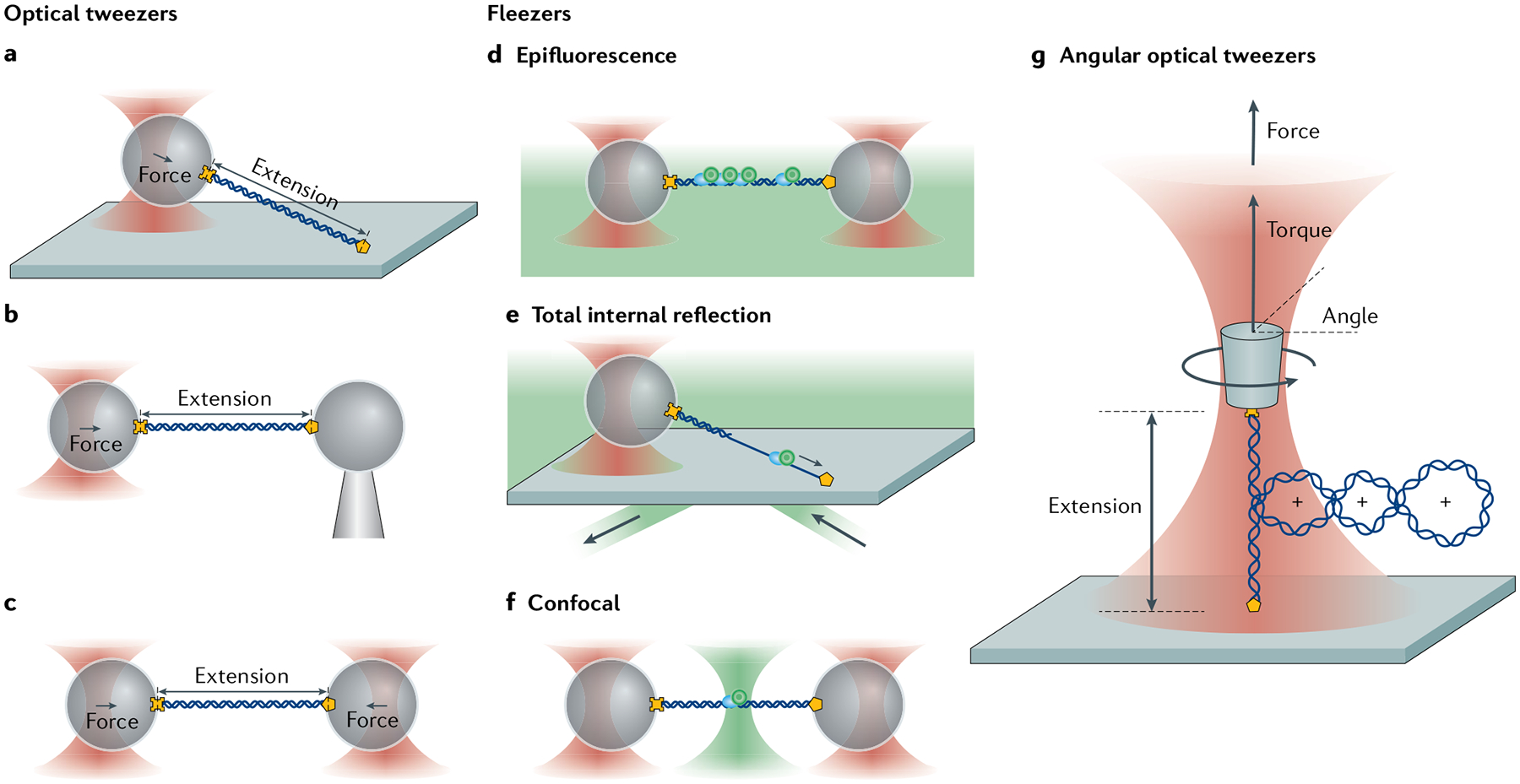
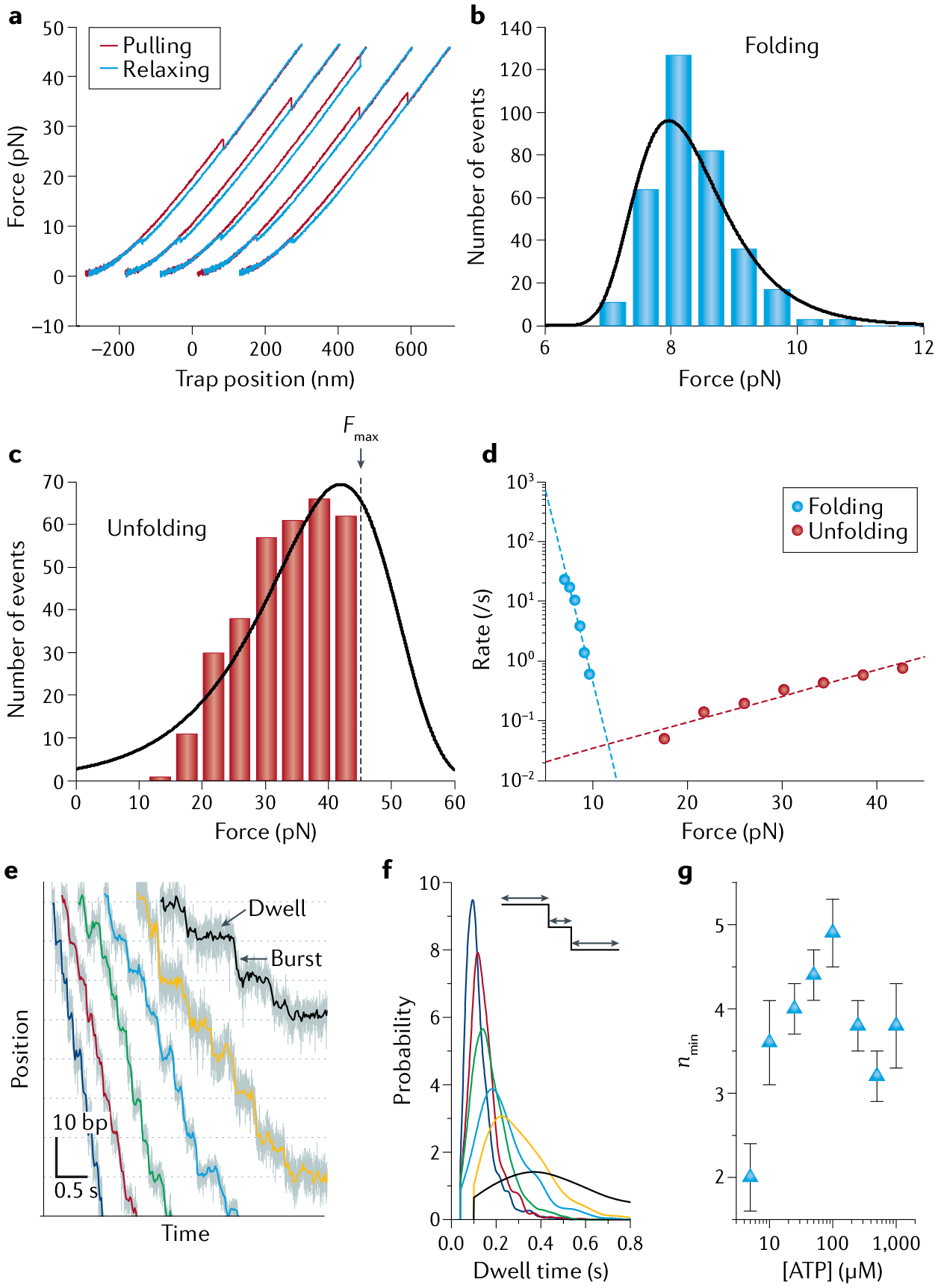
Optical tweezers have become the method of choice in single-molecule manipulation studies. In this Primer, we first review the physical principles of optical tweezers and the characteristics that make them a powerful tool to investigate single molecules. We then introduce the modifications of the method to extend the measurement of forces and displacements to torques and angles, and to develop optical tweezers with single-molecule fluorescence detection capabilities. We discuss force and torque calibration of these instruments, their various modes of operation and most common experimental geometries. We describe the type of data obtained in each experimental design and their analyses. This description is followed by a survey of applications of these methods to the studies of protein-nucleic acid interactions, protein/RNA folding and molecular motors. We also discuss data reproducibility, the factors that lead to the data variability among different laboratories and the need to develop field standards. We cover the current limitations of the methods and possible ways to optimize instrument operation, data extraction and analysis, before suggesting likely areas of future growth.
Conflict of interest statement
Competing interests The authors declare no competing interests.
Figures
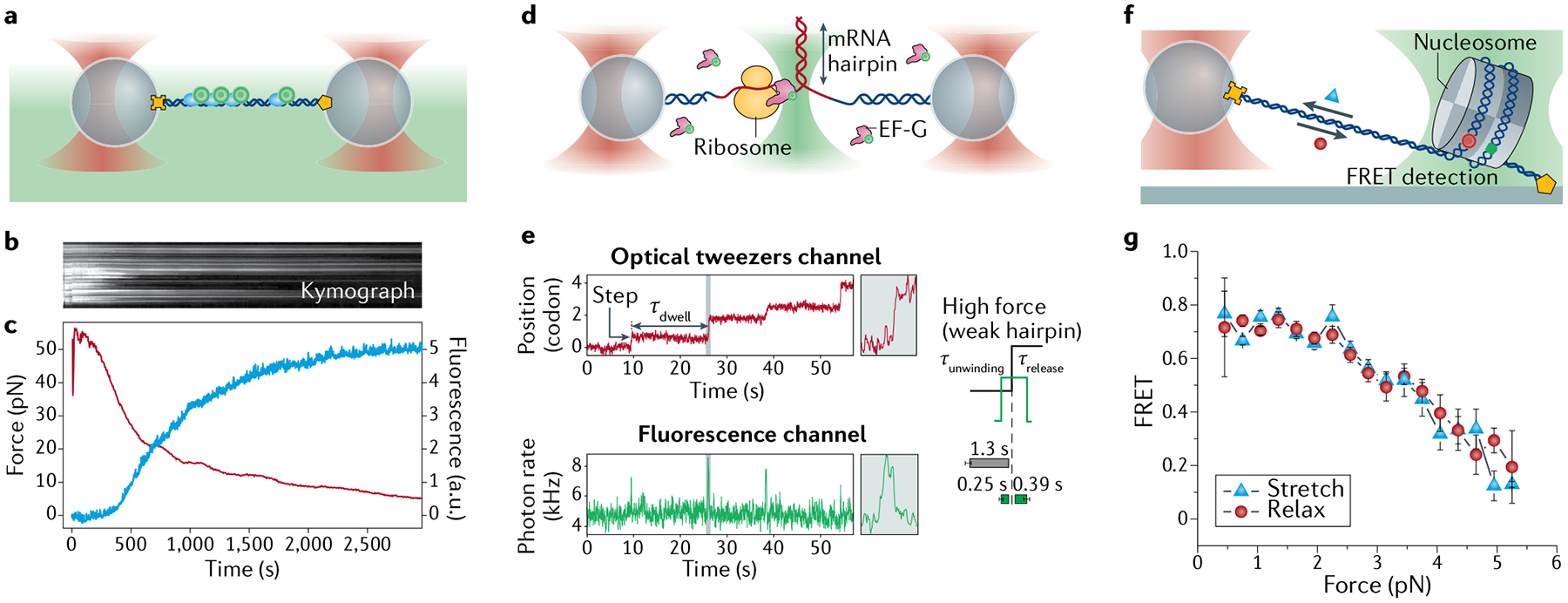
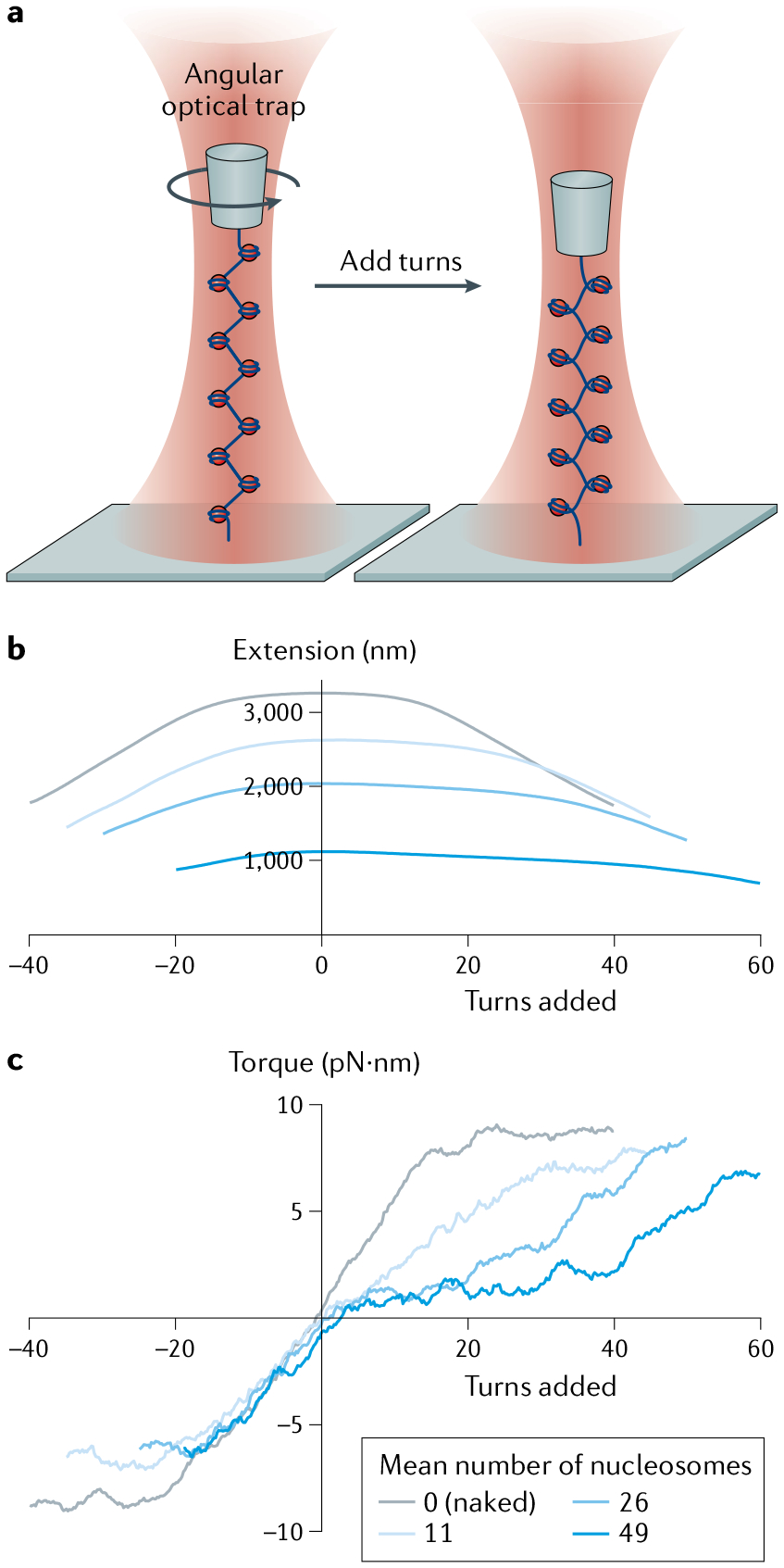
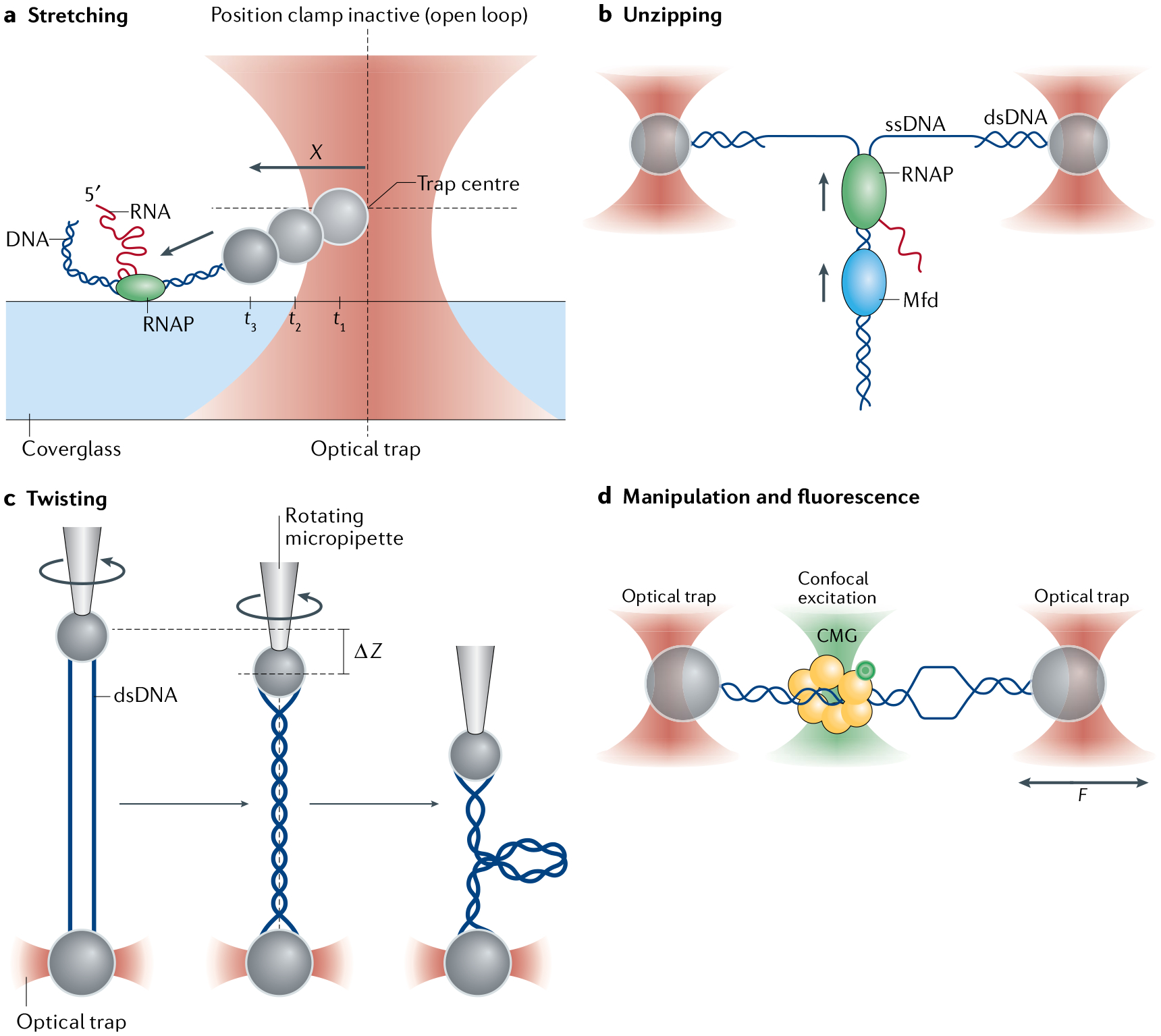
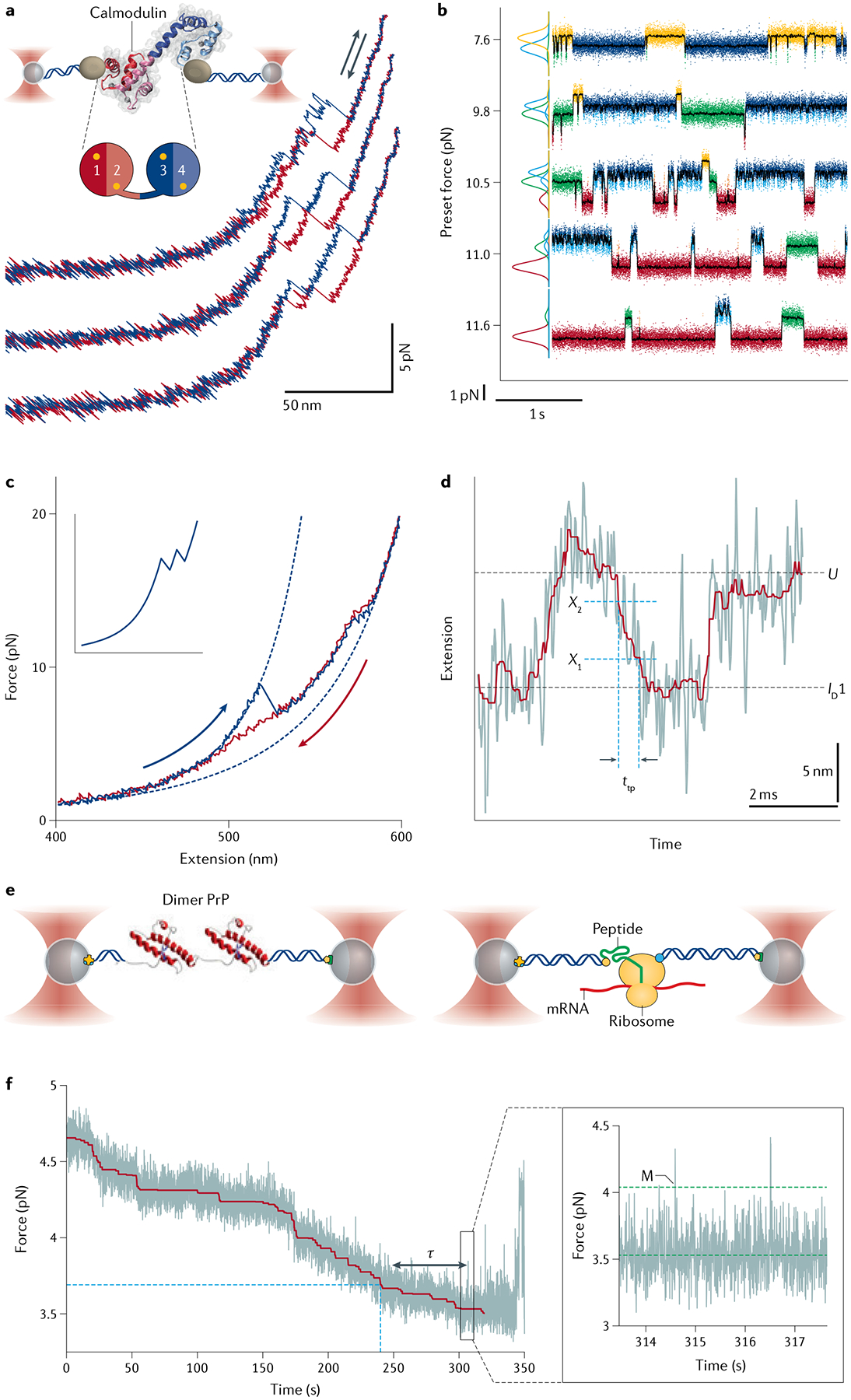
References
-
- Ashkin A Acceleration and trapping of particles by radiation pressure. Phys. Rev. Lett 24, 156 (1970). - PubMed
-
- Ashkin A, Dziedzic JM, Bjorkholm JE & Chu S Observation of a single-beam gradient force optical trap for dielectric particles. Opt. Lett 11, 288 (1986). - PubMed
-
This study provides the first demonstration of a single-beam optical trap that has since been commonly used in single-molecule biophysics.
-
- Ashkin A, Dziedzic JM & Yamane T Optical trapping and manipulation of single cells using infrared laser beams. Nature 330, 769–771 (1987). - PubMed
-
- Jones PH, Marago OM & Volpe G Optical Tweezers: Principles and Applications (Cambridge Univ. Press, 2015).
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources