

Inhibition of SARS-CoV-2 coronavirus proliferation by designer antisense-circRNAs
- PMID: 34850109
- PMCID: PMC8643703
- DOI: 10.1093/nar/gkab1096
Inhibition of SARS-CoV-2 coronavirus proliferation by designer antisense-circRNAs
Abstract
Circular RNAs (circRNAs) are noncoding RNAs that exist in all eukaryotes investigated and are derived from back-splicing of certain pre-mRNA exons. Here, we report the application of artificial circRNAs designed to act as antisense-RNAs. We systematically tested a series of antisense-circRNAs targeted to the SARS-CoV-2 genome RNA, in particular its structurally conserved 5'-untranslated region. Functional assays with both reporter transfections as well as with SARS-CoV-2 infections revealed that specific segments of the SARS-CoV-2 5'-untranslated region can be efficiently accessed by specific antisense-circRNAs, resulting in up to 90% reduction of virus proliferation in cell culture, and with a durability of at least 48 h. Presenting the antisense sequence within a circRNA clearly proved more efficient than in the corresponding linear configuration and is superior to modified antisense oligonucleotides. The activity of the antisense-circRNA is surprisingly robust towards point mutations in the target sequence. This strategy opens up novel applications for designer circRNAs and promising therapeutic strategies in molecular medicine.
© The Author(s) 2021. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
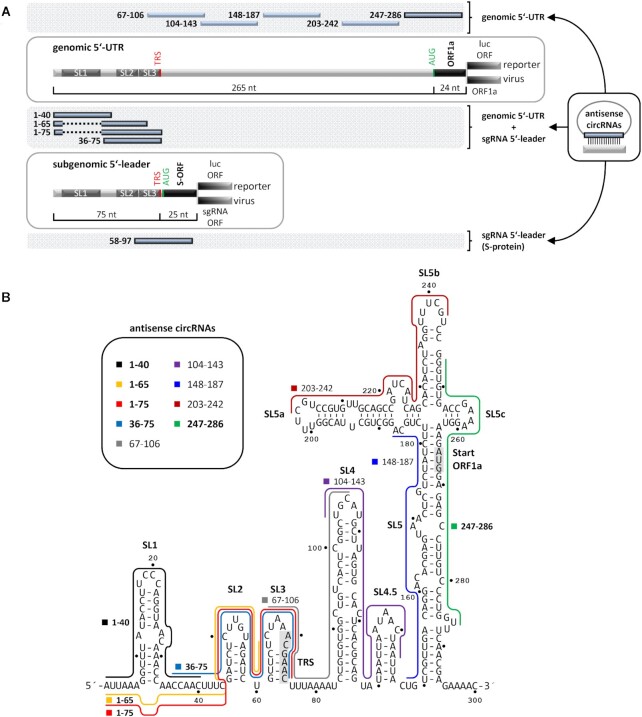
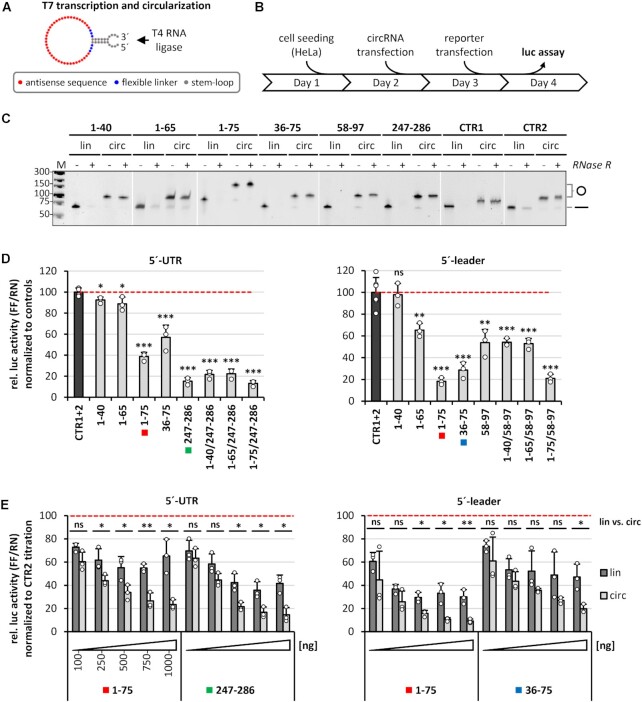
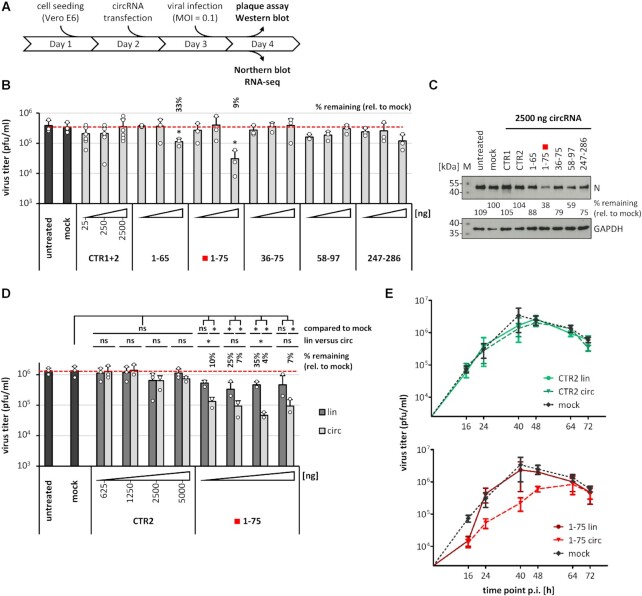
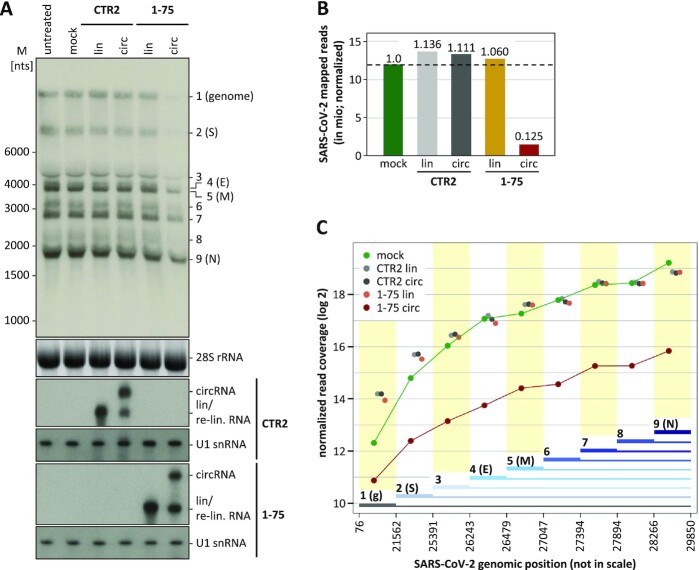
Similar articles
-
Genome-wide mapping of SARS-CoV-2 RNA structures identifies therapeutically-relevant elements.Nucleic Acids Res. 2020 Dec 16;48(22):12436-12452. doi: 10.1093/nar/gkaa1053. Nucleic Acids Res. 2020. PMID: 33166999 Free PMC article.
-
Cryo-EM and antisense targeting of the 28-kDa frameshift stimulation element from the SARS-CoV-2 RNA genome.Nat Struct Mol Biol. 2021 Sep;28(9):747-754. doi: 10.1038/s41594-021-00653-y. Epub 2021 Aug 23. Nat Struct Mol Biol. 2021. PMID: 34426697 Free PMC article.
-
Requirement of the N-terminal region of nonstructural protein 1 in cis for SARS-CoV-2 defective RNA replication.J Virol. 2024 Sep 17;98(9):e0090024. doi: 10.1128/jvi.00900-24. Epub 2024 Aug 28. J Virol. 2024. PMID: 39194239 Free PMC article.
-
Illuminating the pathogenic role of SARS-CoV-2: Insights into competing endogenous RNAs (ceRNAs) regulatory networks.Infect Genet Evol. 2024 Aug;122:105613. doi: 10.1016/j.meegid.2024.105613. Epub 2024 Jun 4. Infect Genet Evol. 2024. PMID: 38844190 Review.
-
Structure and regulation of coronavirus genomes: state-of-the-art and novel insights from SARS-CoV-2 studies.Biochem Soc Trans. 2021 Feb 26;49(1):341-352. doi: 10.1042/BST20200670. Biochem Soc Trans. 2021. PMID: 33367597 Free PMC article. Review.
Cited by
-
Therapeutic prospects of ceRNAs in COVID-19.Front Cell Infect Microbiol. 2022 Sep 20;12:998748. doi: 10.3389/fcimb.2022.998748. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 36204652 Free PMC article. Review.
-
New insights on circular RNAs and their potential applications as biomarkers, therapeutic agents, and preventive vaccines in viral infections: with a glance at SARS-CoV-2.Mol Ther Nucleic Acids. 2022 Sep 13;29:705-717. doi: 10.1016/j.omtn.2022.08.012. Epub 2022 Aug 14. Mol Ther Nucleic Acids. 2022. PMID: 35992045 Free PMC article. Review.
-
Functional Involvement of circRNAs in the Innate Immune Responses to Viral Infection.Viruses. 2023 Aug 5;15(8):1697. doi: 10.3390/v15081697. Viruses. 2023. PMID: 37632040 Free PMC article. Review.
-
The regulation of lncRNAs and miRNAs in SARS-CoV-2 infection.Front Cell Dev Biol. 2023 Jul 27;11:1229393. doi: 10.3389/fcell.2023.1229393. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37576600 Free PMC article. Review.
-
Unlocking the potential of RNA-based therapeutics in the lung: current status and future directions.Front Genet. 2023 Nov 23;14:1281538. doi: 10.3389/fgene.2023.1281538. eCollection 2023. Front Genet. 2023. PMID: 38075698 Free PMC article. Review.
References
-
- Perlman S., Masters P.S.. Howley P.M., Knipe D.M., Whelan S.. Coronaviridae: the viruses and their replication. Fields Virology. 2021; I:Philadelphia, PA: Wolters Kluwer; 410–448.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
