

The nuclear and cytoplasmic activities of RNA polymerase III, and an evolving transcriptome for surveillance
- PMID: 34850129
- PMCID: PMC8643620
- DOI: 10.1093/nar/gkab1145
The nuclear and cytoplasmic activities of RNA polymerase III, and an evolving transcriptome for surveillance
Abstract
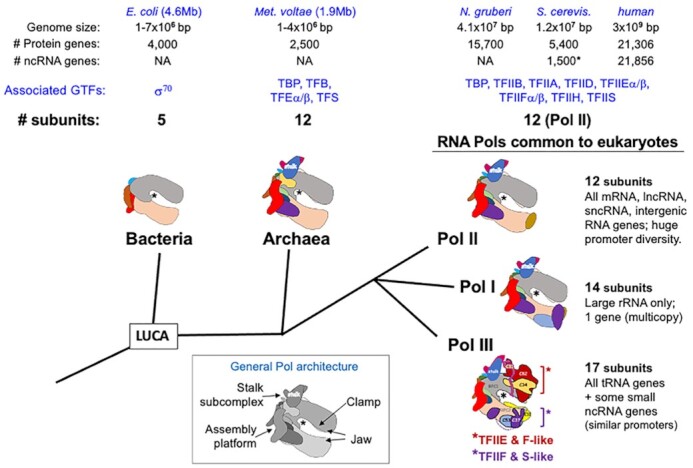
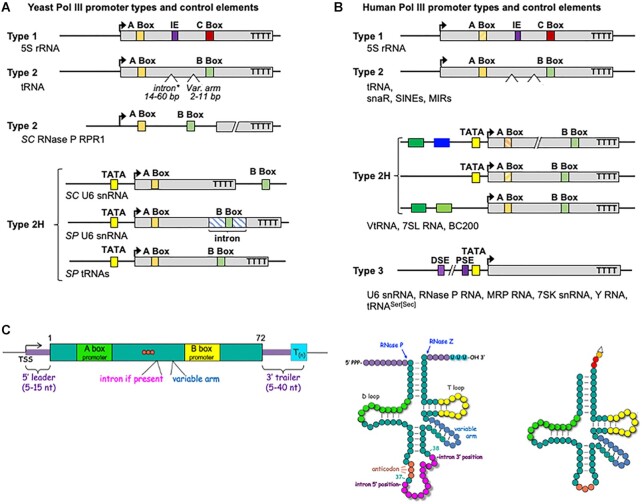
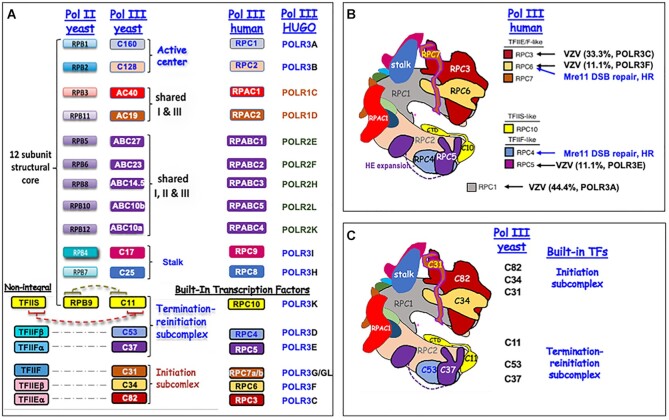
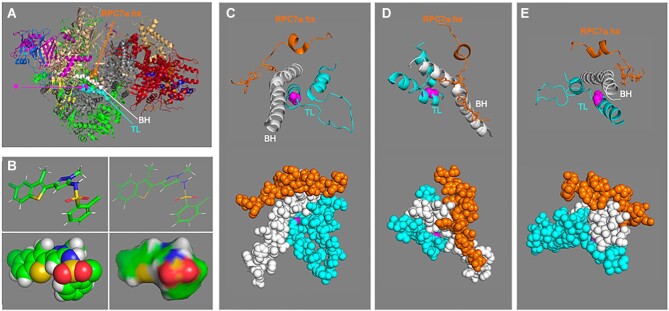
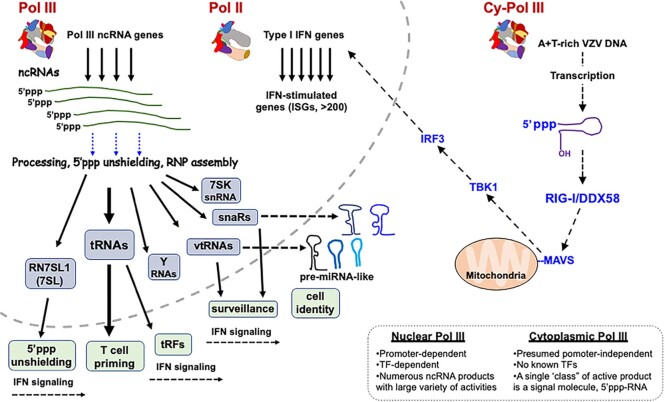
A 1969 report that described biochemical and activity properties of the three eukaryotic RNA polymerases revealed Pol III as highly distinguishable, even before its transcripts were identified. Now known to be the most complex, Pol III contains several stably-associated subunits referred to as built-in transcription factors (BITFs) that enable highly efficient RNA synthesis by a unique termination-associated recycling process. In vertebrates, subunit RPC7(α/β) can be of two forms, encoded by POLR3G or POLR3GL, with differential activity. Here we review promoter-dependent transcription by Pol III as an evolutionary perspective of eukaryotic tRNA expression. Pol III also provides nonconventional functions reportedly by promoter-independent transcription, one of which is RNA synthesis from DNA 3'-ends during repair. Another is synthesis of 5'ppp-RNA signaling molecules from cytoplasmic viral DNA in a pathway of interferon activation that is dysfunctional in immunocompromised patients with mutations in Pol III subunits. These unconventional functions are also reviewed, including evidence that link them to the BITF subunits. We also review data on a fraction of the human Pol III transcriptome that evolved to include vault RNAs and snaRs with activities related to differentiation, and in innate immune and tumor surveillance. The Pol III of higher eukaryotes does considerably more than housekeeping.
Published by Oxford University Press on behalf of Nucleic Acids Research 2021.
Figures
References
-
- Roeder R.G., Rutter W.J.. Multiple forms of DNA-dependent RNA polymerase in eukaryotic organisms. Nature. 1969; 224:234–237. - PubMed
-
- Werner F., Grohmann D. Evolution of multisubunit RNA polymerases in the three domains of life. Nat. Rev. Microbiol. 2011; 9:85–98. - PubMed
-
- Blombach F., Matelska D., Fouqueau T., Cackett G., Werner F.. Key concepts and challenges in archaeal transcription. J. Mol. Biol. 2019; 431:4184–4201. - PubMed
-
- Gurdon J., Brown D.D.. The transcription of 5S DNA injected into xenopus oocytes. Dev. Biol. 1978; 67:346–356. - PubMed
-
- Sklar V.E., Roeder R.G.. Transcription of specific genes in isolated nuclei by exogenous RNA polymerases. Cell. 1977; 10:405–414. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
