

Heterogeneity of human anti-viral immunity shaped by virus, tissue, age, and sex
- PMID: 34852222
- PMCID: PMC8719595
- DOI: 10.1016/j.celrep.2021.110071
Heterogeneity of human anti-viral immunity shaped by virus, tissue, age, and sex
Abstract
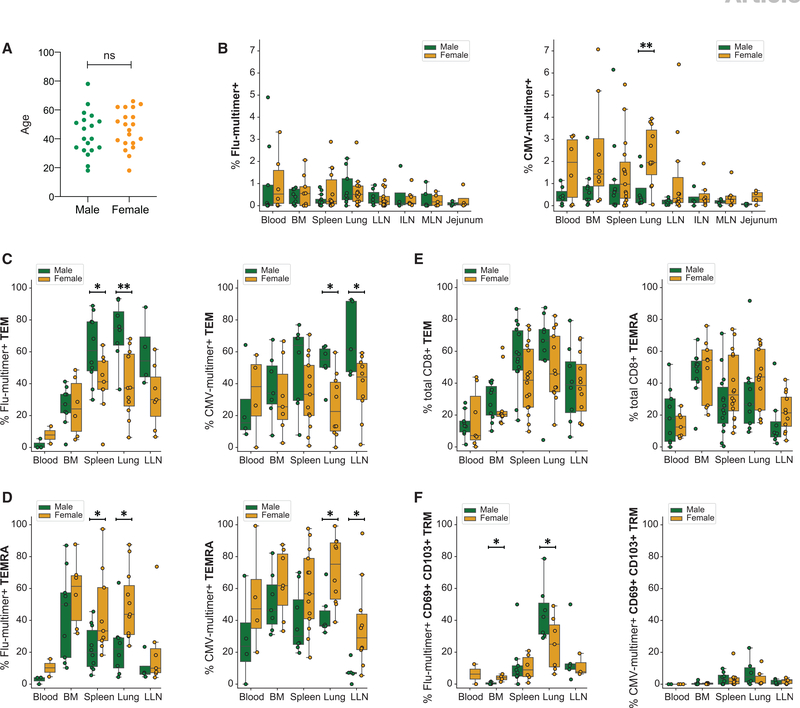
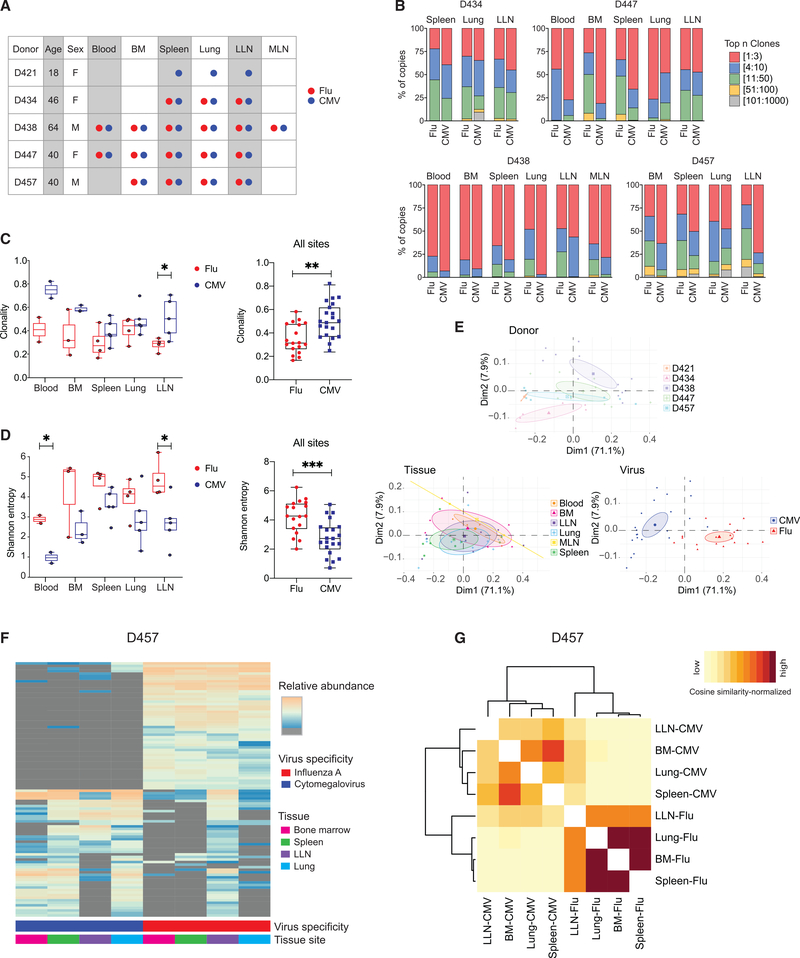
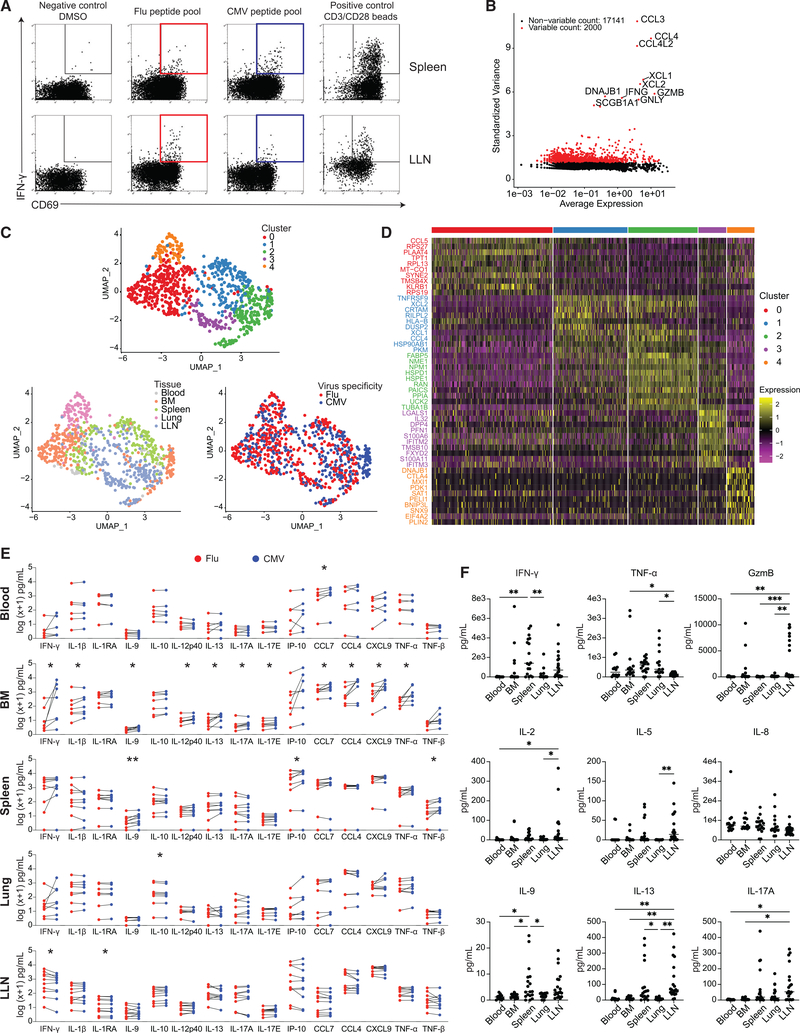
The persistence of anti-viral immunity is essential for protection and exhibits profound heterogeneity across individuals. Here, we elucidate the factors that shape maintenance and function of anti-viral T cell immunity in the body by comprehensive profiling of virus-specific T cells across blood, lymphoid organs, and mucosal tissues of organ donors. We use flow cytometry, T cell receptor sequencing, single-cell transcriptomics, and cytokine analysis to profile virus-specific CD8+ T cells recognizing the ubiquitous pathogens influenza and cytomegalovirus. Our results reveal that virus specificity determines overall magnitude, tissue distribution, differentiation, and clonal repertoire of virus-specific T cells. Age and sex influence T cell differentiation and dissemination in tissues, while T cell tissue residence and functionality are highly correlated with the site. Together, our results demonstrate how the covariates of virus, tissue, age, and sex impact the anti-viral immune response, which is important for targeting, monitoring, and predicting immune responses to existing and emerging viruses.
Keywords: T lymphocytes; anti-viral immunity; cytomegalovirus; human immunology; immunological memory; influenza; lymphoid organs; mucosal immunity; tissue resident memory T cells.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.S. is a consultant for Gritstone, Flow Pharma, Arcturus, Immunoscape, CellCarta, OxfordImmunotech, and Avalia. La Jolla Institute for Immunology (LJI) has filed for patent protection for various aspects of T cell epitope and vaccine design work. E.T.L.P. is an advisor for Roche Diagnostics, Enpicom, The Antibody Society, IEDB, and The American Autoimmune Related Diseases Association. D.L.F. is a consultant for Moderna.
Figures
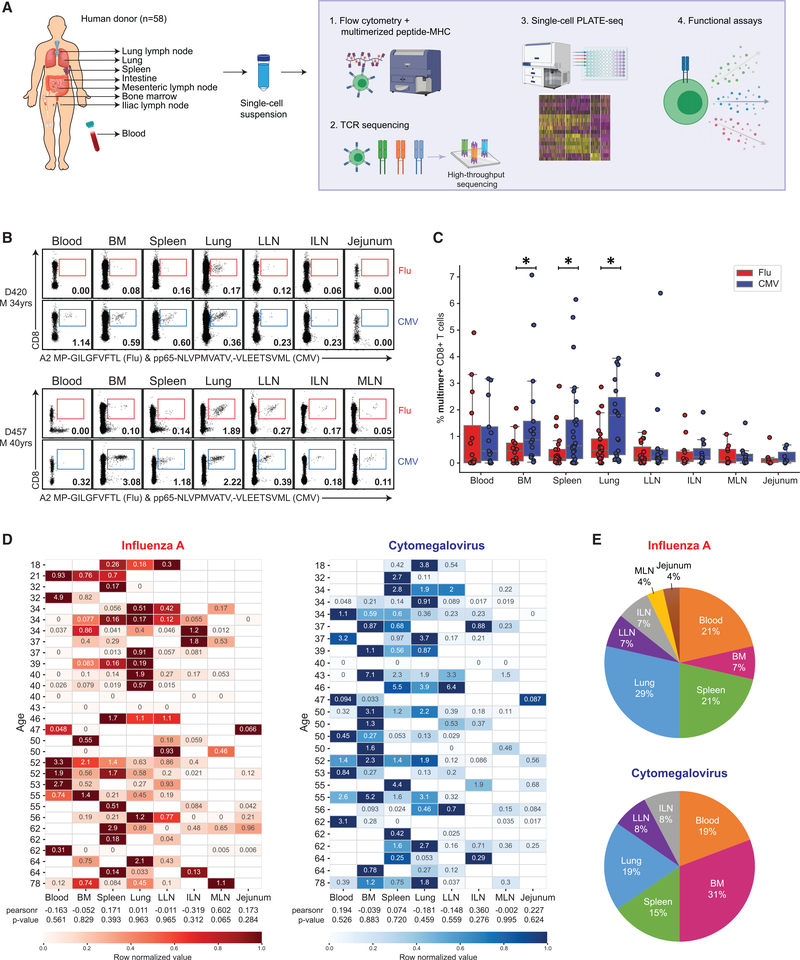
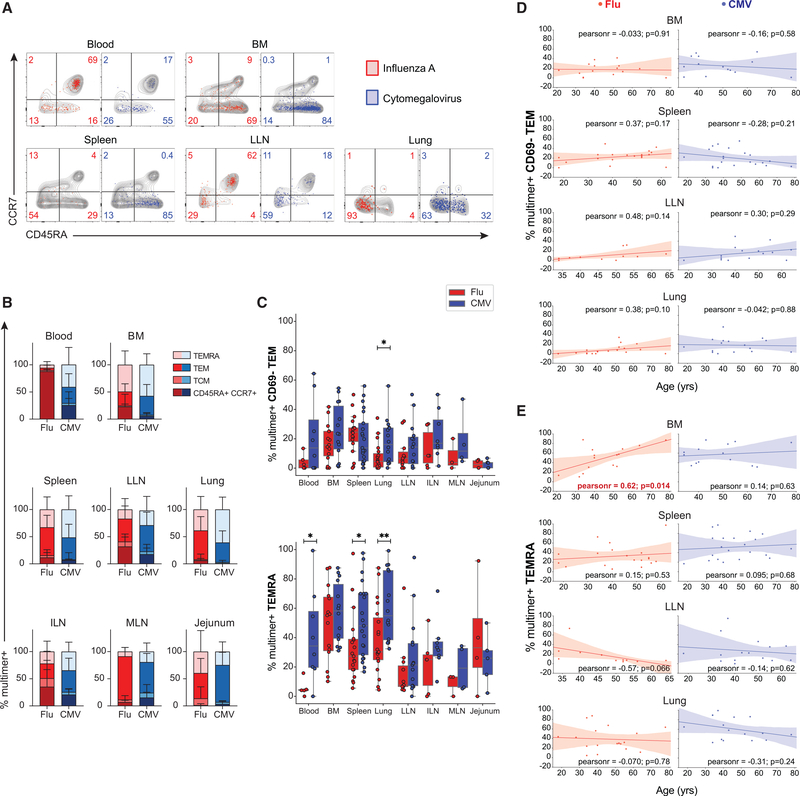
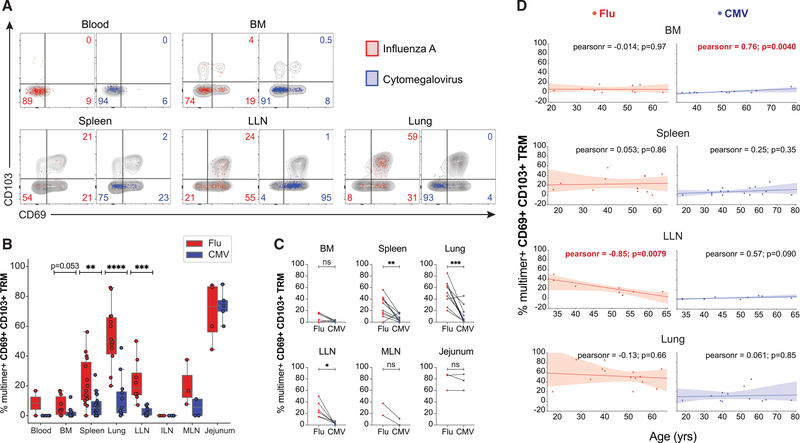
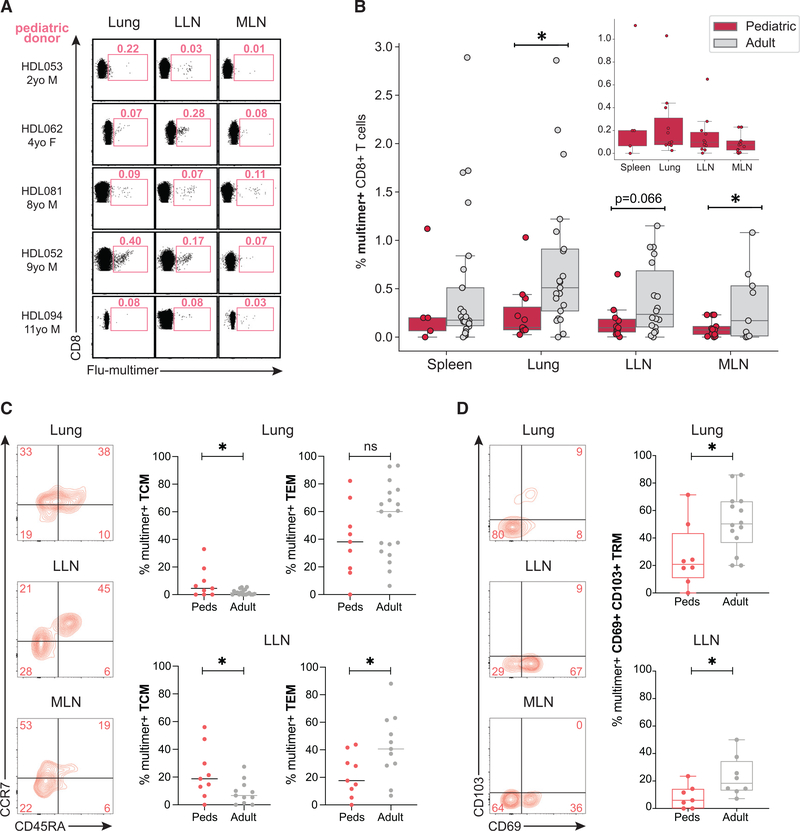
References
-
- Abdullah M, Chai PS, Chong MY, Tohit ER, Ramasamy R, Pei CP, and Vidyadaran S (2012). Gender effect on in vitro lymphocyte subset levels of healthy individuals. Cell. Immunol. 272, 214–219. - PubMed
-
- Bolotin DA, Poslavsky S, Mitrophanov I, Shugay M, Mamedov IZ, Putintseva EV, and Chudakov DM (2015). MiXCR: software for comprehensive adaptive immunity profiling. Nat. Methods 12, 380–381. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
