

Neoantigen-driven B cell and CD4 T follicular helper cell collaboration promotes anti-tumor CD8 T cell responses
- PMID: 34852236
- PMCID: PMC8671355
- DOI: 10.1016/j.cell.2021.11.007
Neoantigen-driven B cell and CD4 T follicular helper cell collaboration promotes anti-tumor CD8 T cell responses
Abstract
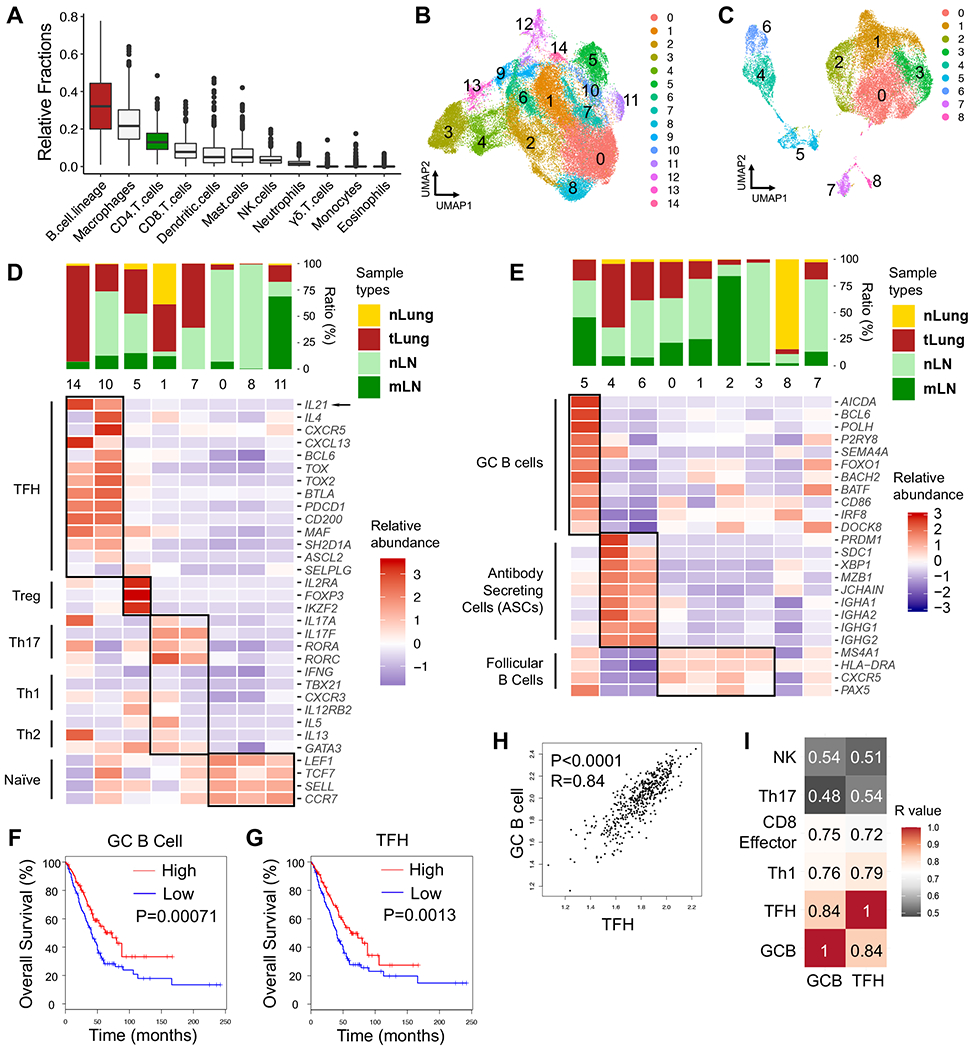
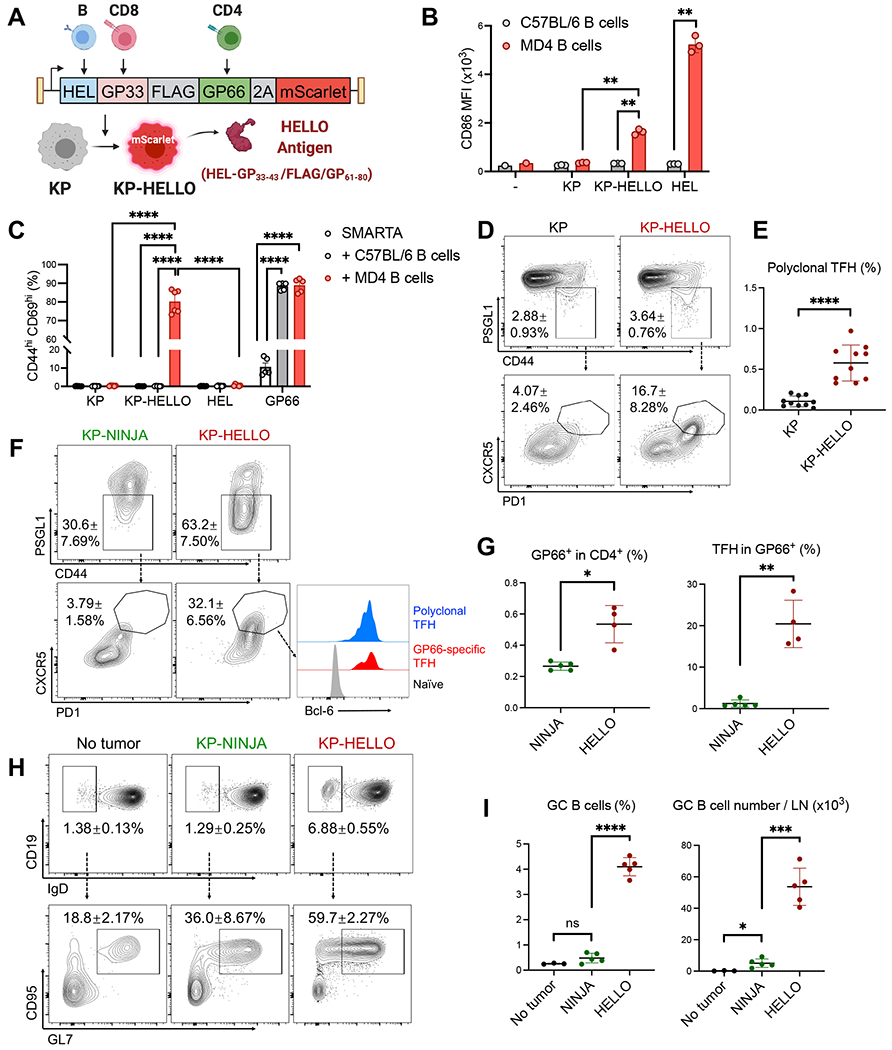
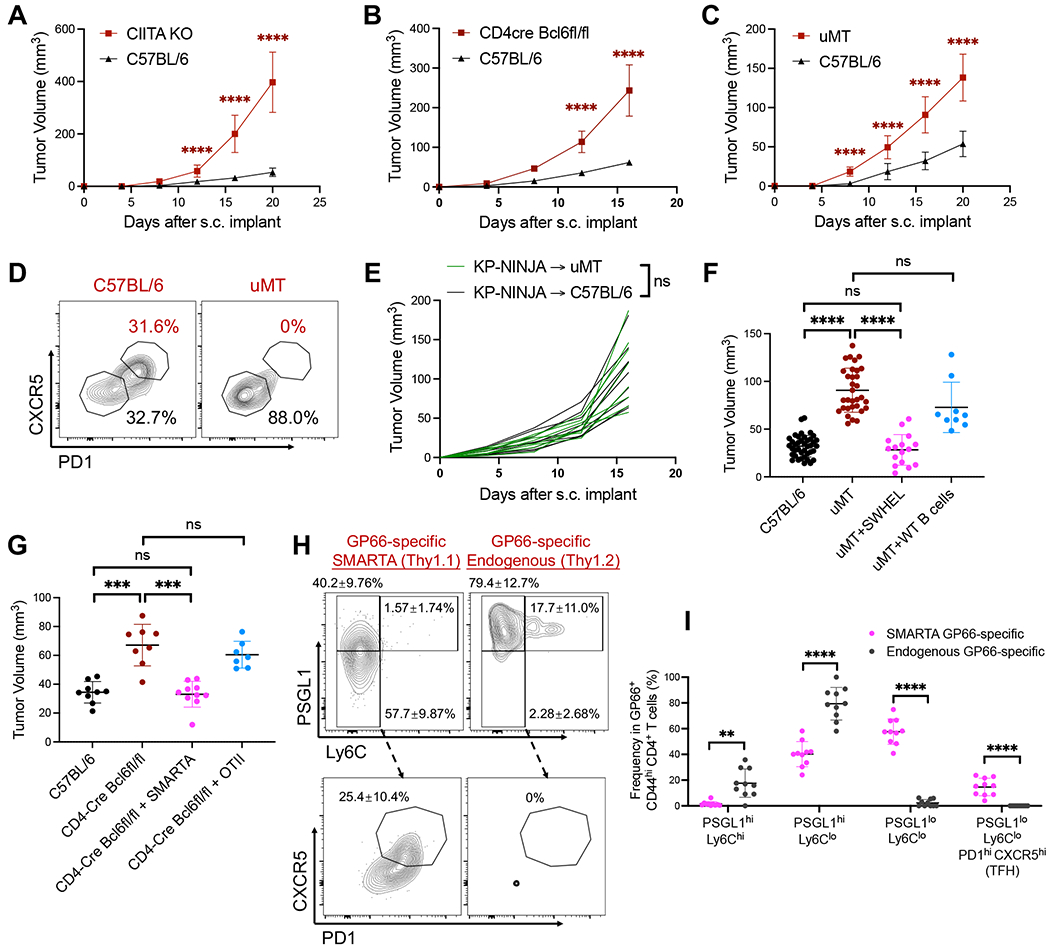
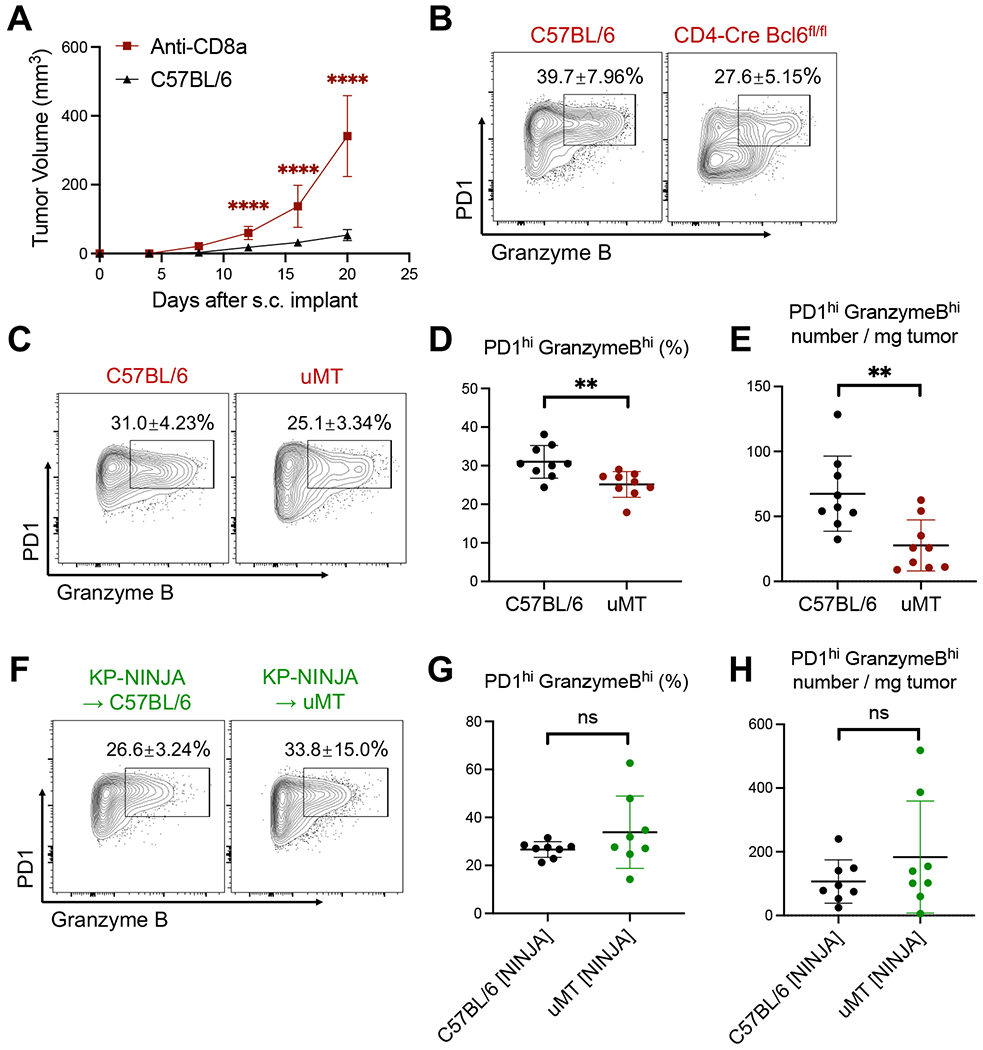
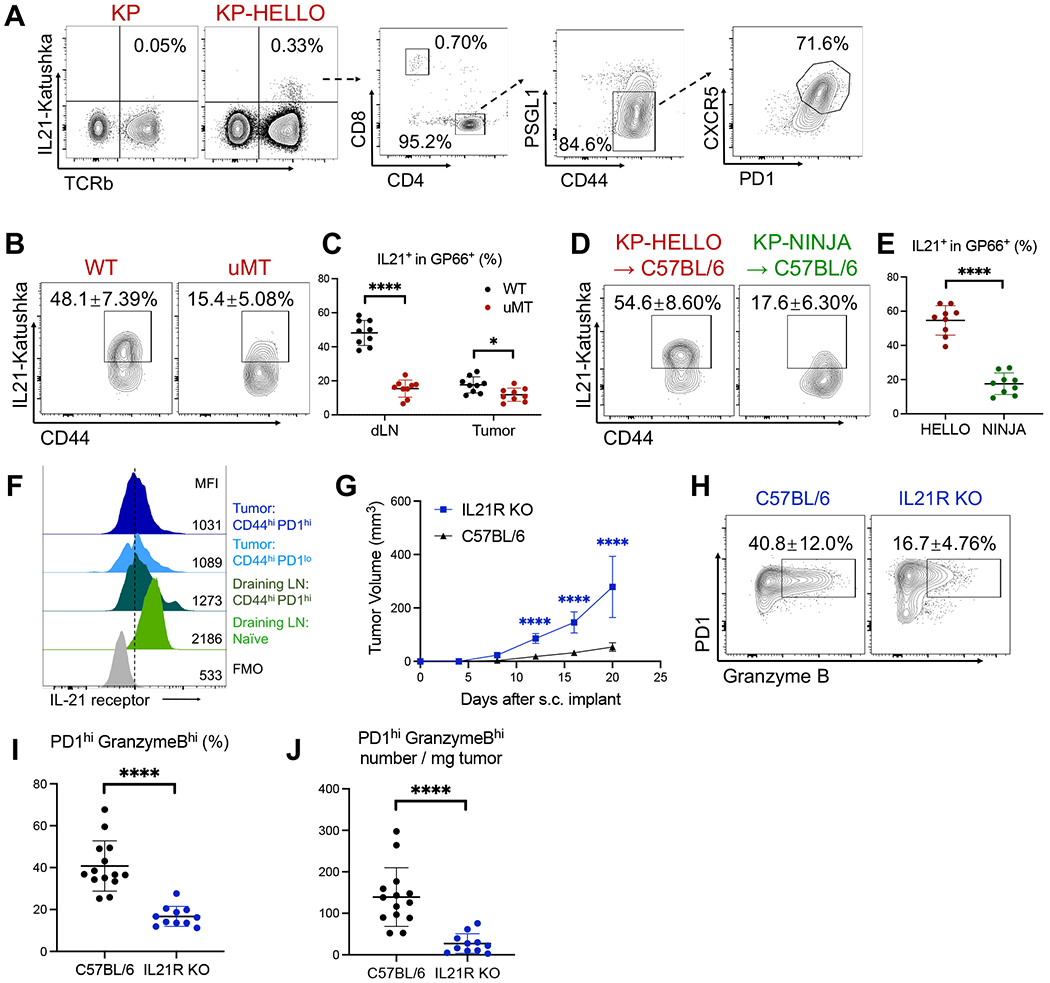
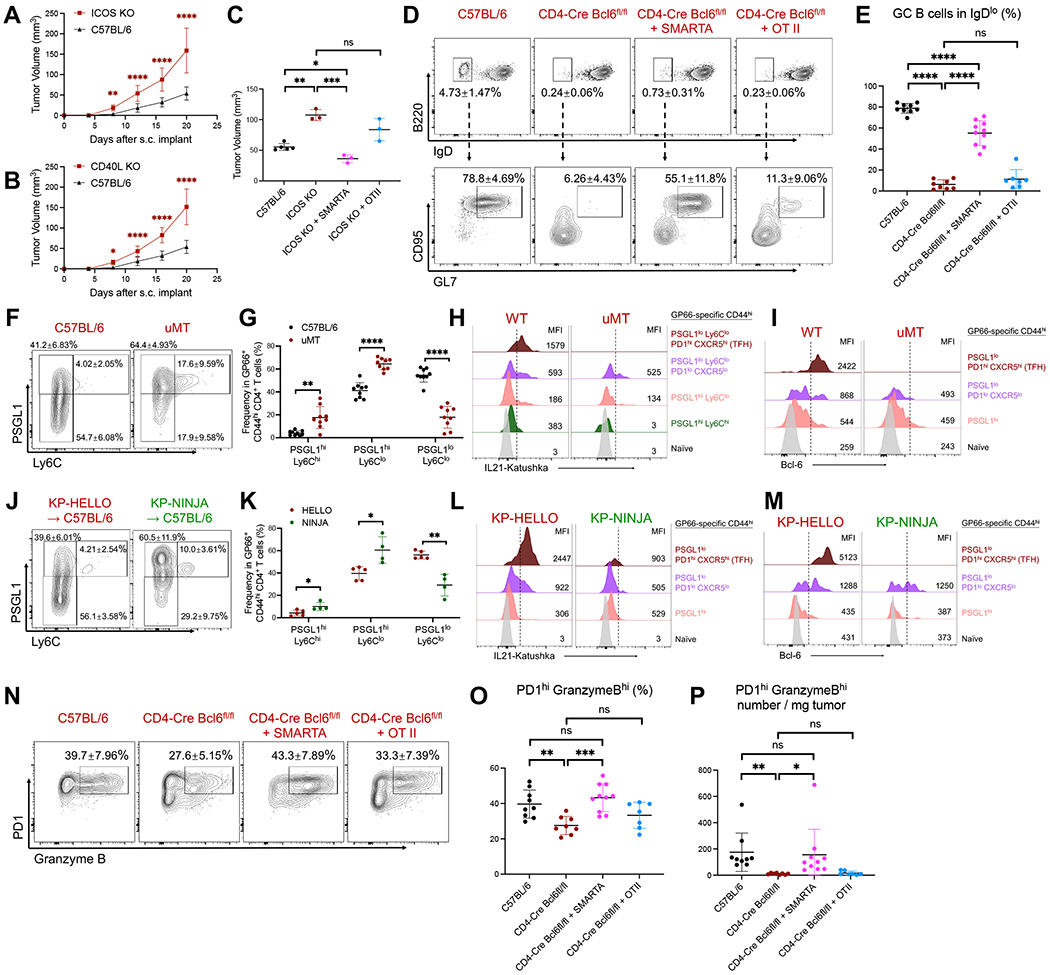
CD4 T follicular helper (TFH) cells support B cells, which are critical for germinal center (GC) formation, but the importance of TFH-B cell interactions in cancer is unclear. We found enrichment of TFH cell transcriptional signature correlates with GC B cell signature and with prolonged survival in individuals with lung adenocarcinoma (LUAD). We further developed a murine LUAD model in which tumor cells express B cell- and T cell-recognized neoantigens. Interactions between tumor-specific TFH and GC B cells, as well as interleukin (IL)-21 primarily produced by TFH cells, are necessary for tumor control and effector CD8 T cell function. Development of TFH cells requires B cells and B cell-recognized neoantigens. Thus, tumor neoantigens can regulate the fate of tumor-specific CD4 T cells by facilitating their interactions with tumor-specific B cells, which in turn promote anti-tumor immunity by enhancing CD8 T cell effector functions.
Keywords: B cell; CD8 T cell; IL-21; T follicular helper cell; lung cancer; neoantigen.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
