

Histone variant H2A.Z regulates zygotic genome activation
- PMID: 34853314
- PMCID: PMC8636486
- DOI: 10.1038/s41467-021-27125-7
Histone variant H2A.Z regulates zygotic genome activation
Abstract
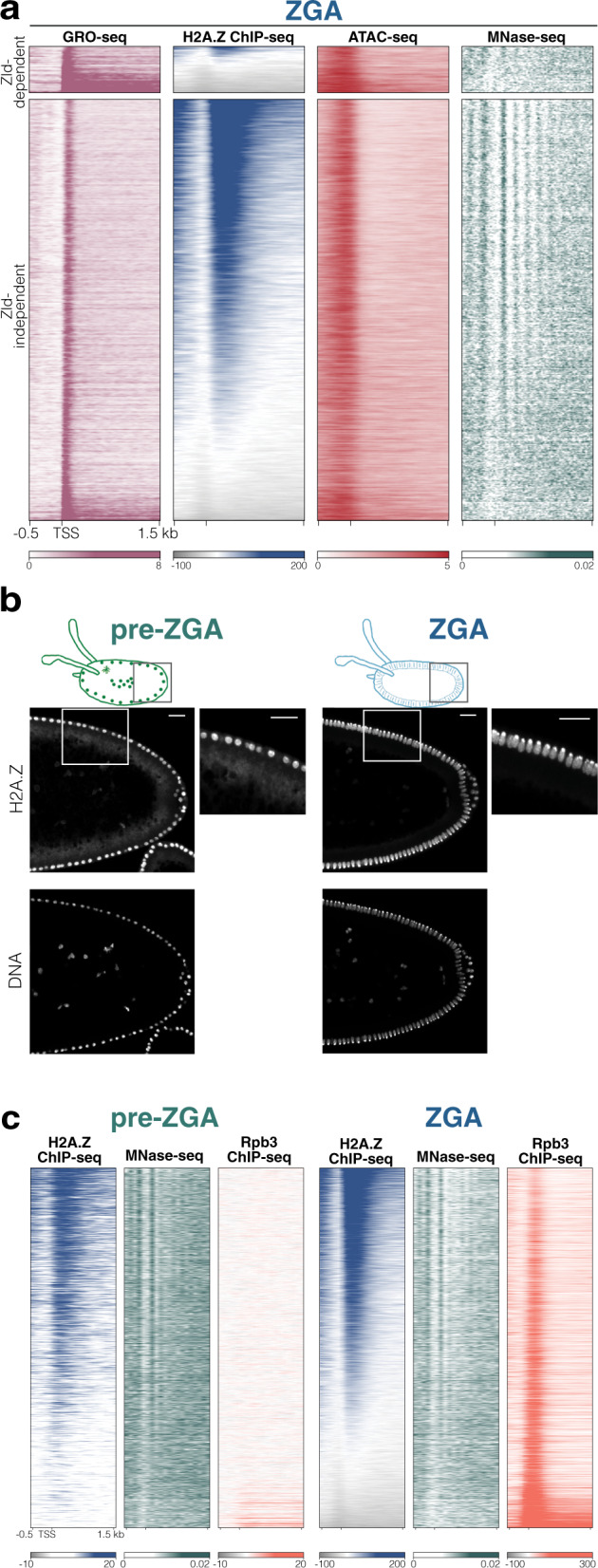
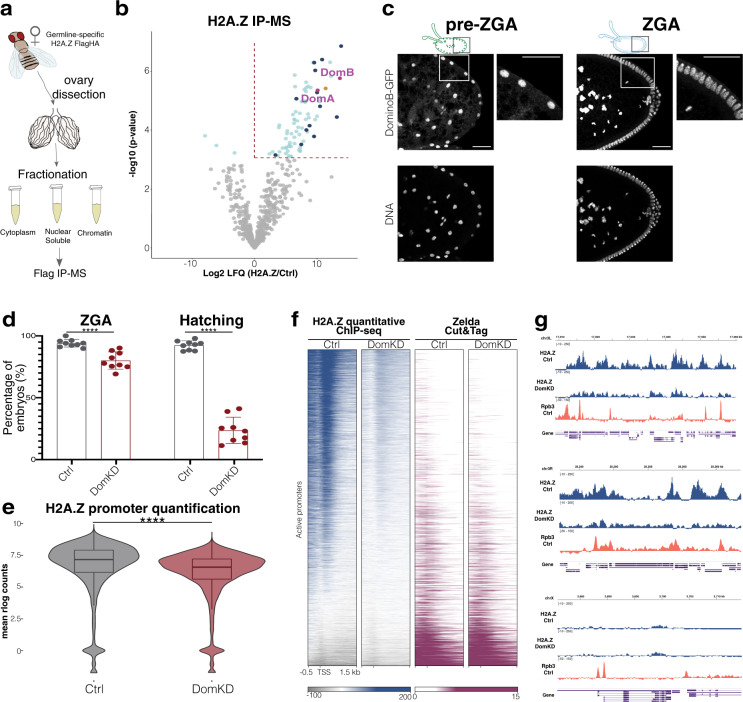
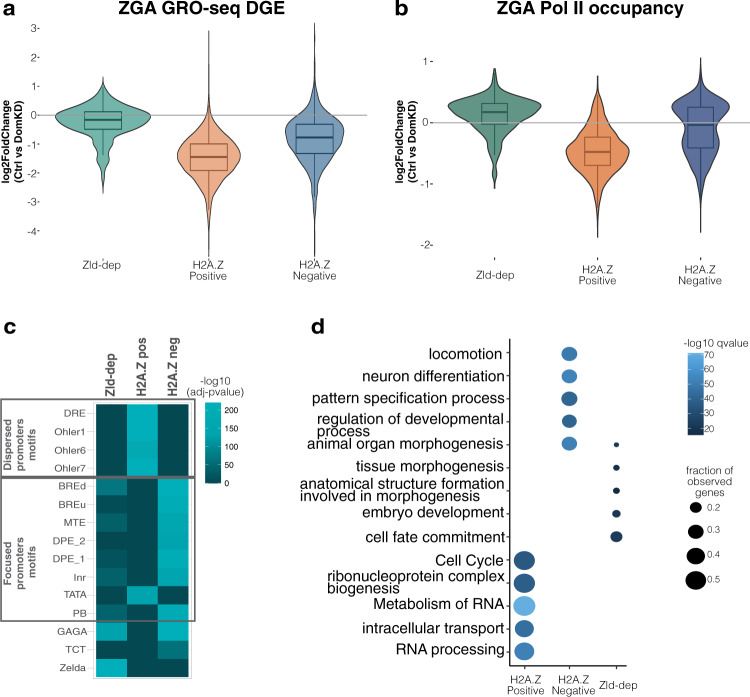
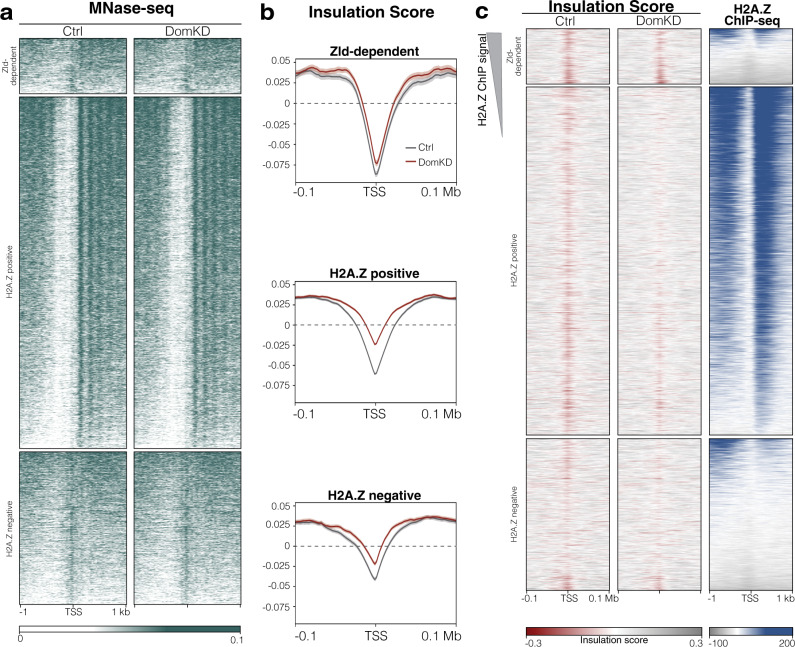
During embryogenesis, the genome shifts from transcriptionally quiescent to extensively active in a process known as Zygotic Genome Activation (ZGA). In Drosophila, the pioneer factor Zelda is known to be essential for the progression of development; still, it regulates the activation of only a small subset of genes at ZGA. However, thousands of genes do not require Zelda, suggesting that other mechanisms exist. By conducting GRO-seq, HiC and ChIP-seq in Drosophila embryos, we demonstrate that up to 65% of zygotically activated genes are enriched for the histone variant H2A.Z. H2A.Z enrichment precedes ZGA and RNA Polymerase II loading onto chromatin. In vivo knockdown of maternally contributed Domino, a histone chaperone and ATPase, reduces H2A.Z deposition at transcription start sites, causes global downregulation of housekeeping genes at ZGA, and compromises the establishment of the 3D chromatin structure. We infer that H2A.Z is essential for the de novo establishment of transcriptional programs during ZGA via chromatin reorganization.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Exchanges of histone methylation and variants during mouse zygotic genome activation.Zygote. 2020 Feb;28(1):51-58. doi: 10.1017/S0967199419000649. Epub 2019 Nov 20. Zygote. 2020. PMID: 31746724
-
Histone concentration regulates the cell cycle and transcription in early development.Development. 2019 Oct 4;146(19):dev177402. doi: 10.1242/dev.177402. Development. 2019. PMID: 31511251 Free PMC article.
-
CLAMP regulates zygotic genome activation in Drosophila embryos.Genetics. 2021 Oct 2;219(2):iyab107. doi: 10.1093/genetics/iyab107. Genetics. 2021. PMID: 34849887 Free PMC article.
-
Epigenetic inheritance and gene expression regulation in early Drosophila embryos.EMBO Rep. 2024 Oct;25(10):4131-4152. doi: 10.1038/s44319-024-00245-z. Epub 2024 Sep 16. EMBO Rep. 2024. PMID: 39285248 Free PMC article. Review.
-
The emergence of genome architecture and zygotic genome activation.Curr Opin Cell Biol. 2020 Jun;64:50-57. doi: 10.1016/j.ceb.2020.02.002. Epub 2020 Mar 19. Curr Opin Cell Biol. 2020. PMID: 32220807 Free PMC article. Review.
Cited by
-
The Function of H2A Histone Variants and Their Roles in Diseases.Biomolecules. 2024 Aug 12;14(8):993. doi: 10.3390/biom14080993. Biomolecules. 2024. PMID: 39199381 Free PMC article. Review.
-
The Drosophila embryo as a tabula rasa for the epigenome.Fac Rev. 2022 Dec 23;11:40. doi: 10.12703/r/11-40. eCollection 2022. Fac Rev. 2022. PMID: 36644296 Free PMC article. Review.
-
Setting the stage for development: the maternal-to-zygotic transition in Drosophila.Genetics. 2023 Oct 4;225(2):iyad142. doi: 10.1093/genetics/iyad142. Genetics. 2023. PMID: 37616526 Free PMC article. Review.
-
The maternal-to-zygotic transition: reprogramming of the cytoplasm and nucleus.Nat Rev Genet. 2025 Apr;26(4):245-267. doi: 10.1038/s41576-024-00792-0. Epub 2024 Nov 25. Nat Rev Genet. 2025. PMID: 39587307 Free PMC article. Review.
-
IMPACTS OF DNA METHYLATION ON H2A.Z DEPOSITION AND NUCLEOSOME STABILITY.bioRxiv [Preprint]. 2025 Jul 31:2025.07.31.667981. doi: 10.1101/2025.07.31.667981. bioRxiv. 2025. PMID: 40766494 Free PMC article. Preprint.
References
-
- Vastenhouw, N. L., Cao, W. X. & Lipshitz, H. D. The maternal-to-zygotic transition revisited. Development146, dev161471 (2019). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
