

Local circuit amplification of spatial selectivity in the hippocampus
- PMID: 34853473
- PMCID: PMC9746172
- DOI: 10.1038/s41586-021-04169-9
Local circuit amplification of spatial selectivity in the hippocampus
Abstract
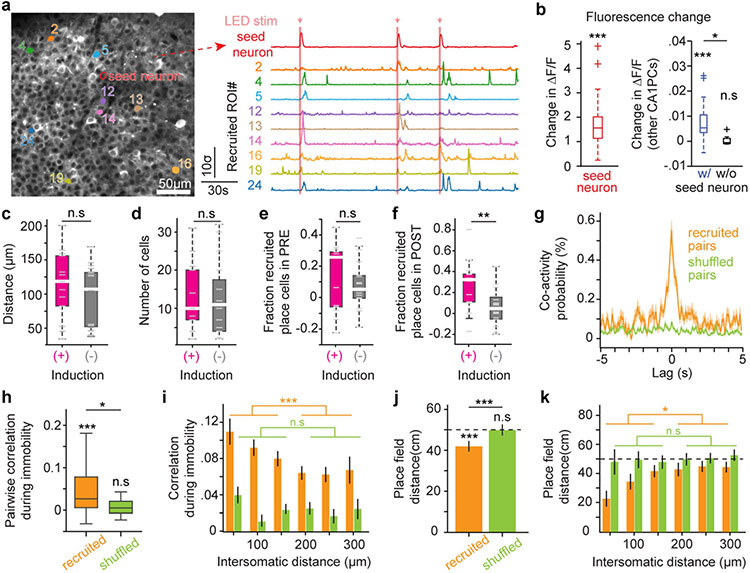
Local circuit architecture facilitates the emergence of feature selectivity in the cerebral cortex1. In the hippocampus, it remains unknown whether local computations supported by specific connectivity motifs2 regulate the spatial receptive fields of pyramidal cells3. Here we developed an in vivo electroporation method for monosynaptic retrograde tracing4 and optogenetics manipulation at single-cell resolution to interrogate the dynamic interaction of place cells with their microcircuitry during navigation. We found a local circuit mechanism in CA1 whereby the spatial tuning of an individual place cell can propagate to a functionally recurrent subnetwork5 to which it belongs. The emergence of place fields in individual neurons led to the development of inverse selectivity in a subset of their presynaptic interneurons, and recruited functionally coupled place cells at that location. Thus, the spatial selectivity of single CA1 neurons is amplified through local circuit plasticity to enable effective multi-neuronal representations that can flexibly scale environmental features locally without degrading the feedforward input structure.
© 2021. The Author(s), under exclusive licence to Springer Nature Limited.
Figures













References
-
- O’Keefe J & Dostrovsky J The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Res. 34, 171–175 (1971). - PubMed
-
- Wertz A et al. Single-cell-initiated monosynaptic tracing reveals layer-specific cortical network modules. Science 349, 70–74 (2015). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
