

Fumarate is a terminal electron acceptor in the mammalian electron transport chain
- PMID: 34855504
- PMCID: PMC8803114
- DOI: 10.1126/science.abi7495
Fumarate is a terminal electron acceptor in the mammalian electron transport chain
Abstract
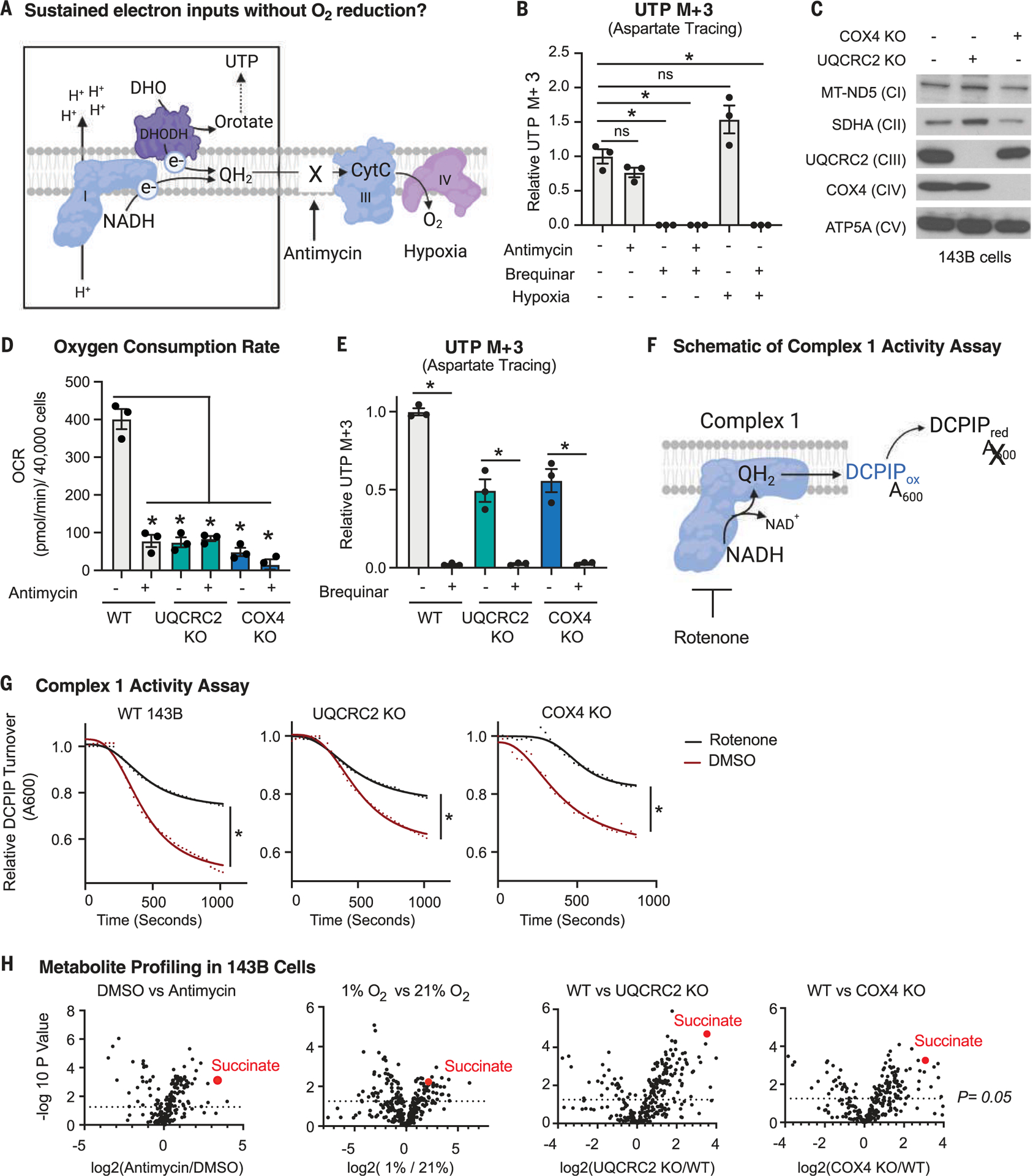
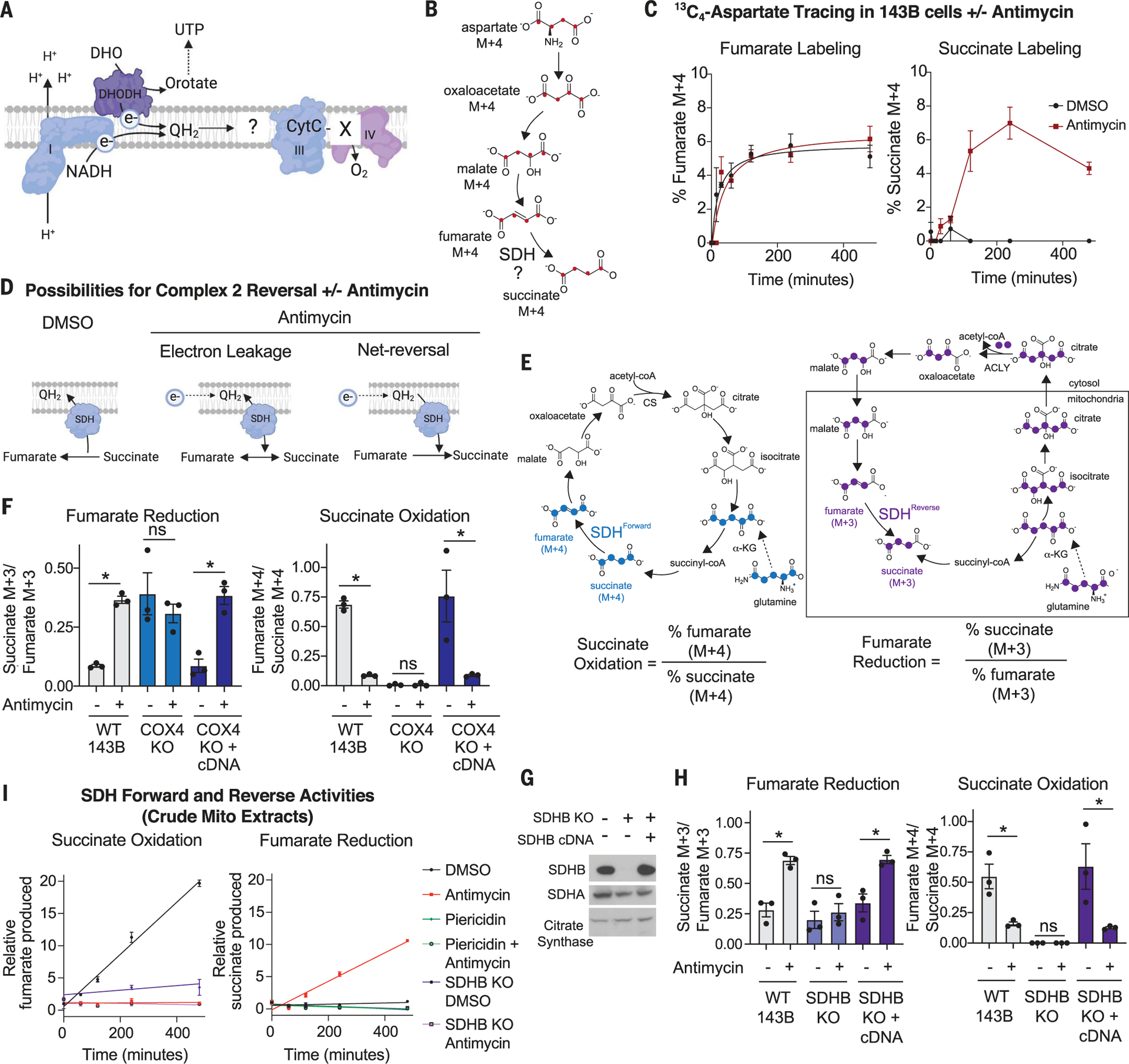
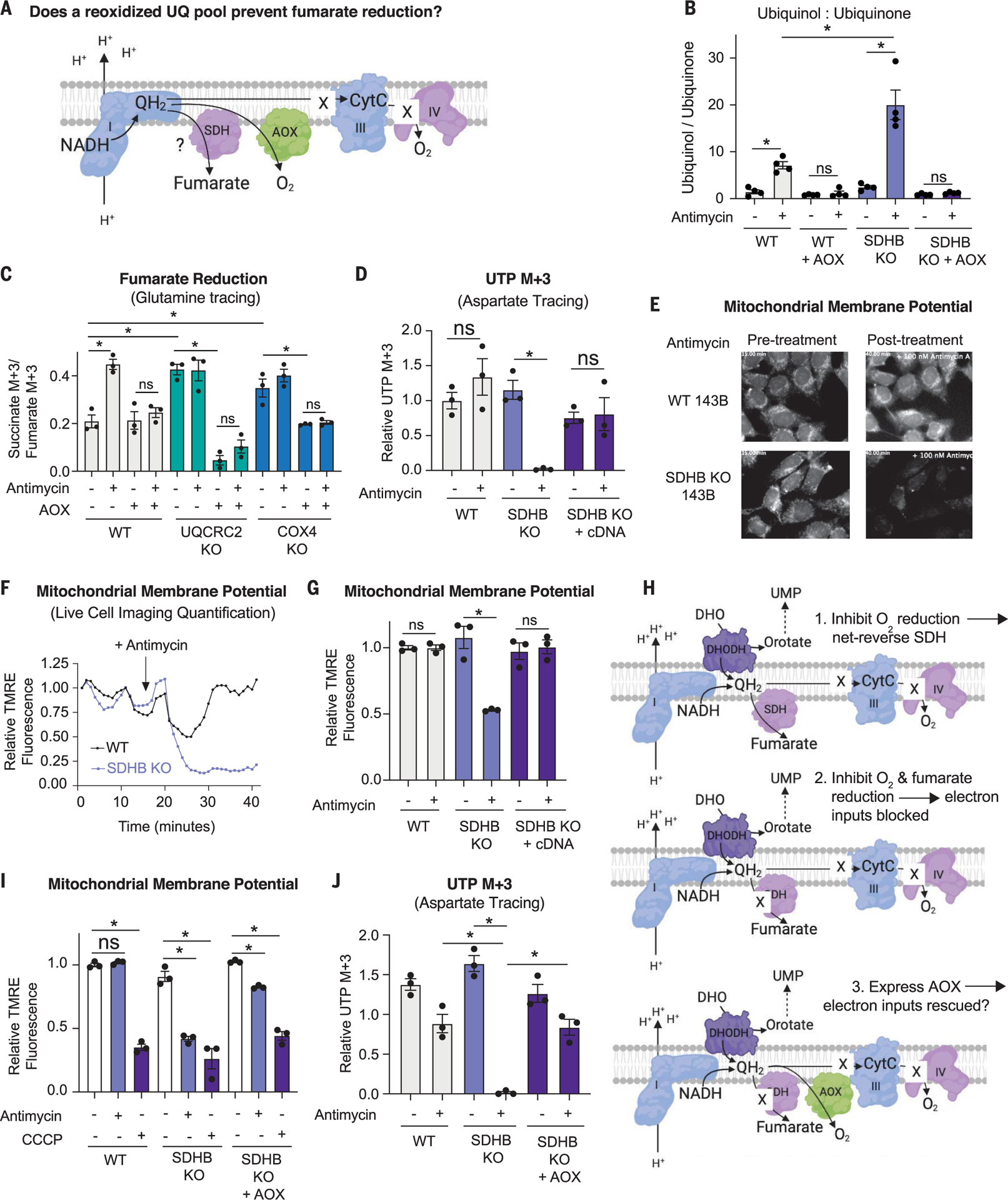
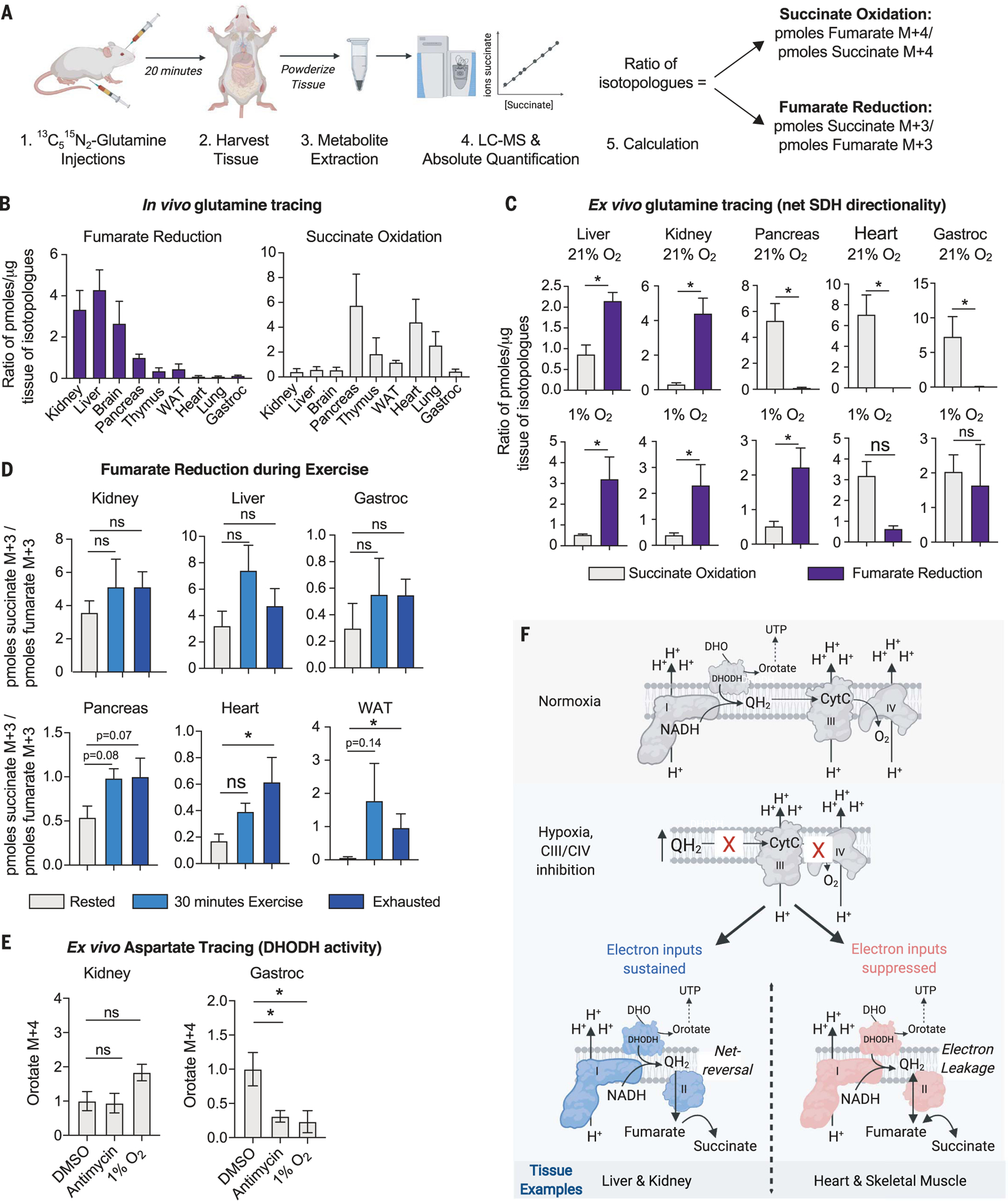
For electrons to continuously enter and flow through the mitochondrial electron transport chain (ETC), they must ultimately land on a terminal electron acceptor (TEA), which is known to be oxygen in mammals. Paradoxically, we find that complex I and dihydroorotate dehydrogenase (DHODH) can still deposit electrons into the ETC when oxygen reduction is impeded. Cells lacking oxygen reduction accumulate ubiquinol, driving the succinate dehydrogenase (SDH) complex in reverse to enable electron deposition onto fumarate. Upon inhibition of oxygen reduction, fumarate reduction sustains DHODH and complex I activities. Mouse tissues display varying capacities to use fumarate as a TEA, most of which net reverse the SDH complex under hypoxia. Thus, we delineate a circuit of electron flow in the mammalian ETC that maintains mitochondrial functions under oxygen limitation.
Conflict of interest statement
Figures
Comment in
-
Short-circuiting respiration.Science. 2021 Dec 3;374(6572):1196-1197. doi: 10.1126/science.abm8098. Epub 2021 Dec 2. Science. 2021. PMID: 34855498
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
