

Bis-choline tetrathiomolybdate prevents copper-induced blood-brain barrier damage
- PMID: 34857647
- PMCID: PMC8675913
- DOI: 10.26508/lsa.202101164
Bis-choline tetrathiomolybdate prevents copper-induced blood-brain barrier damage
Abstract
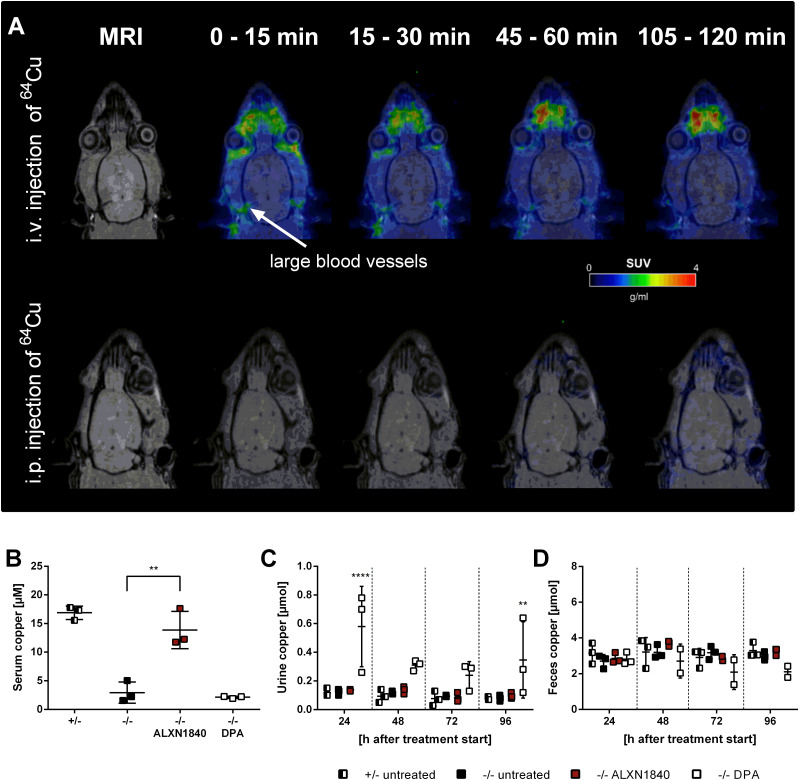
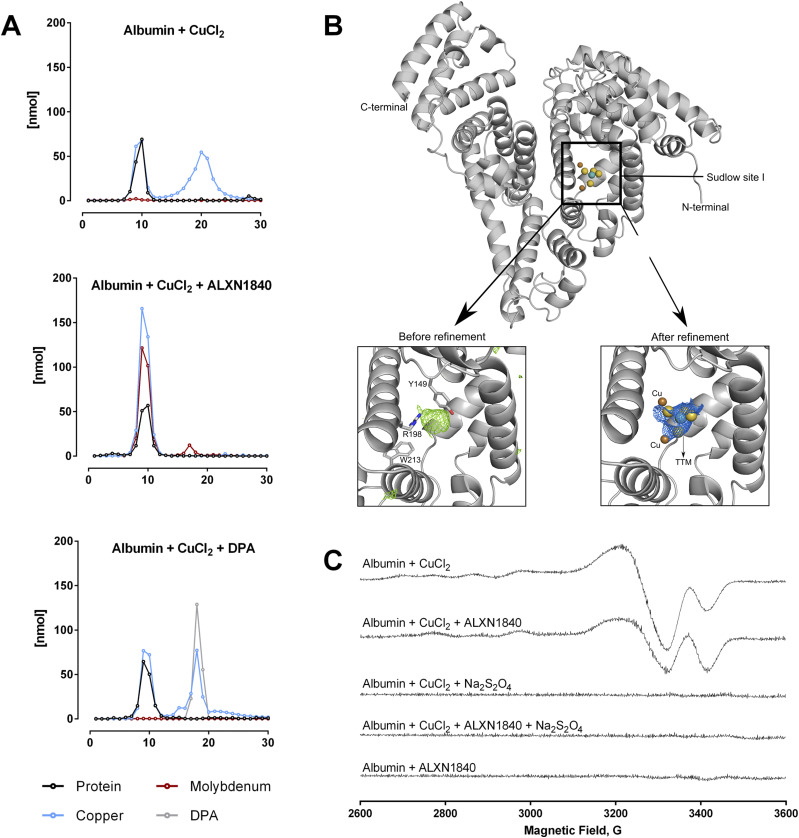
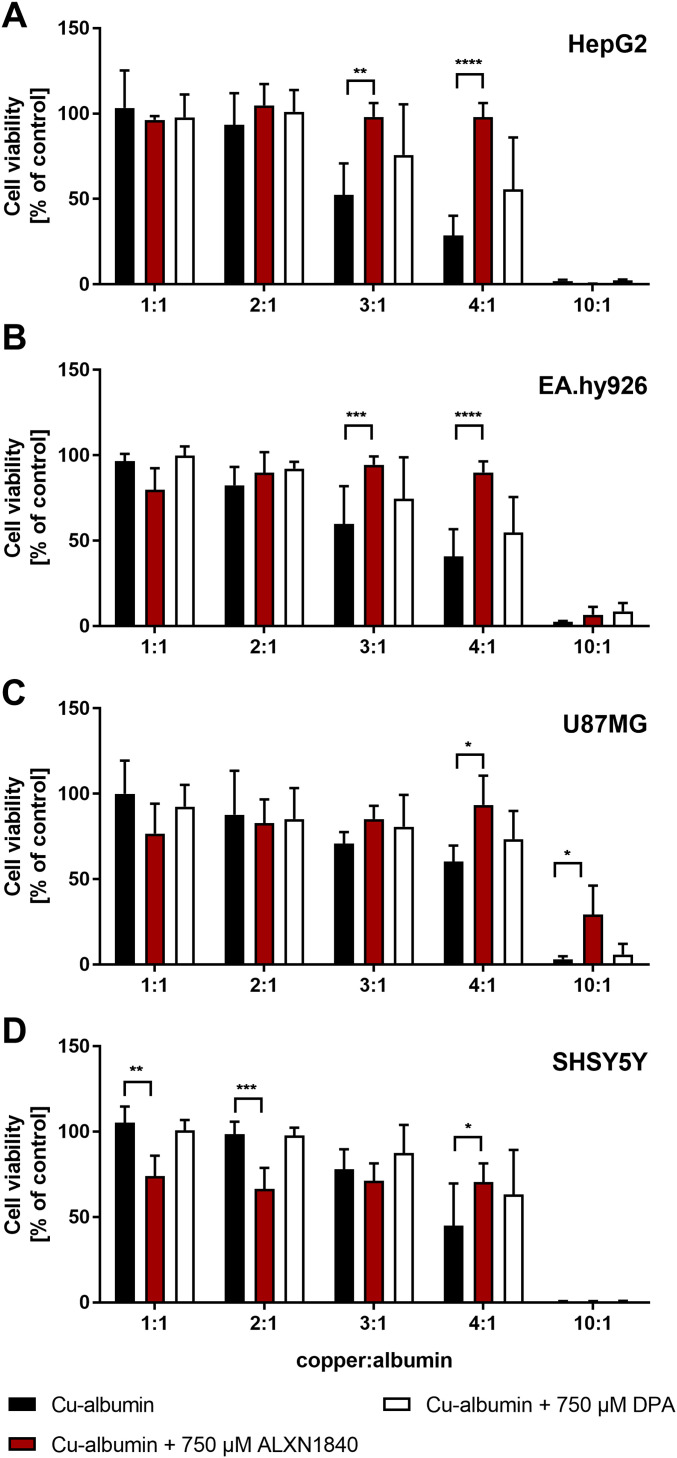
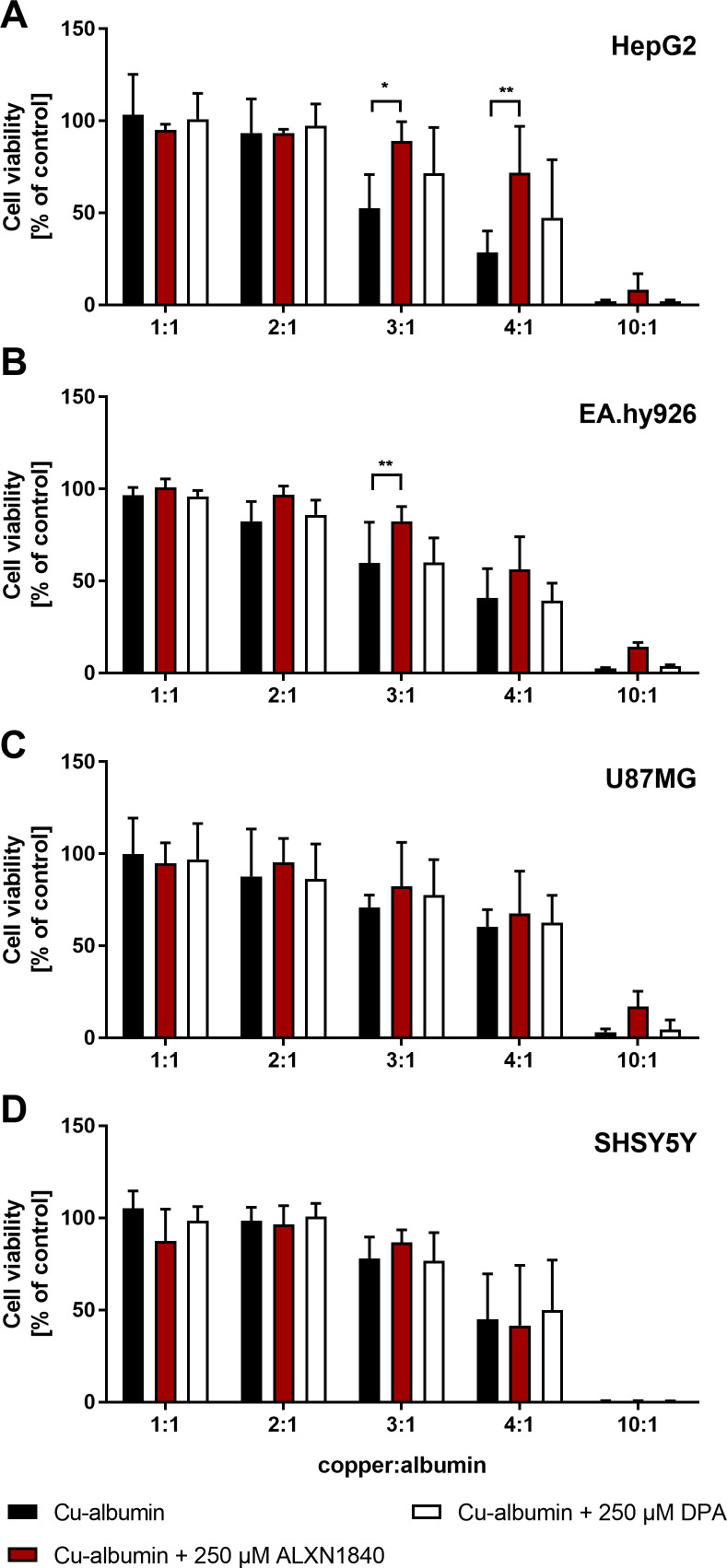
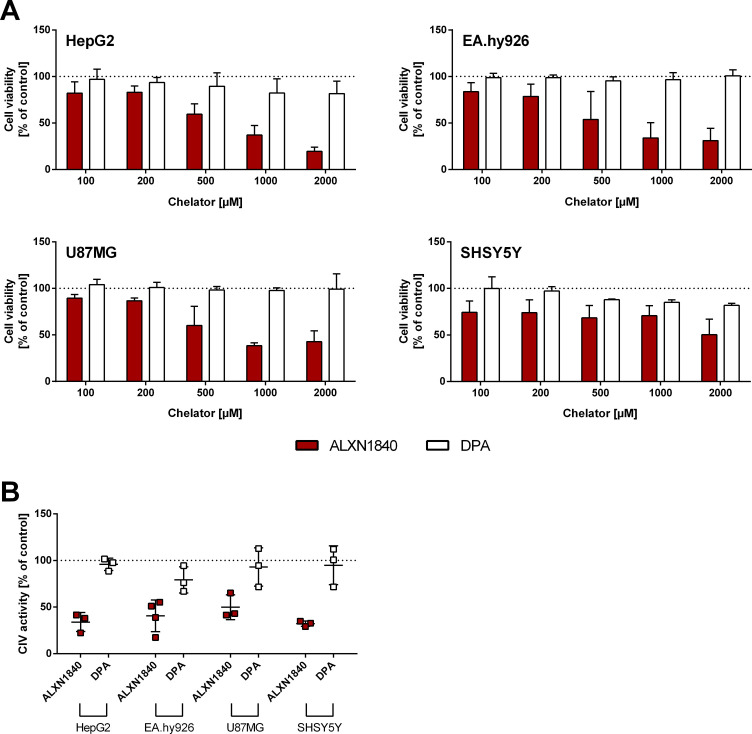
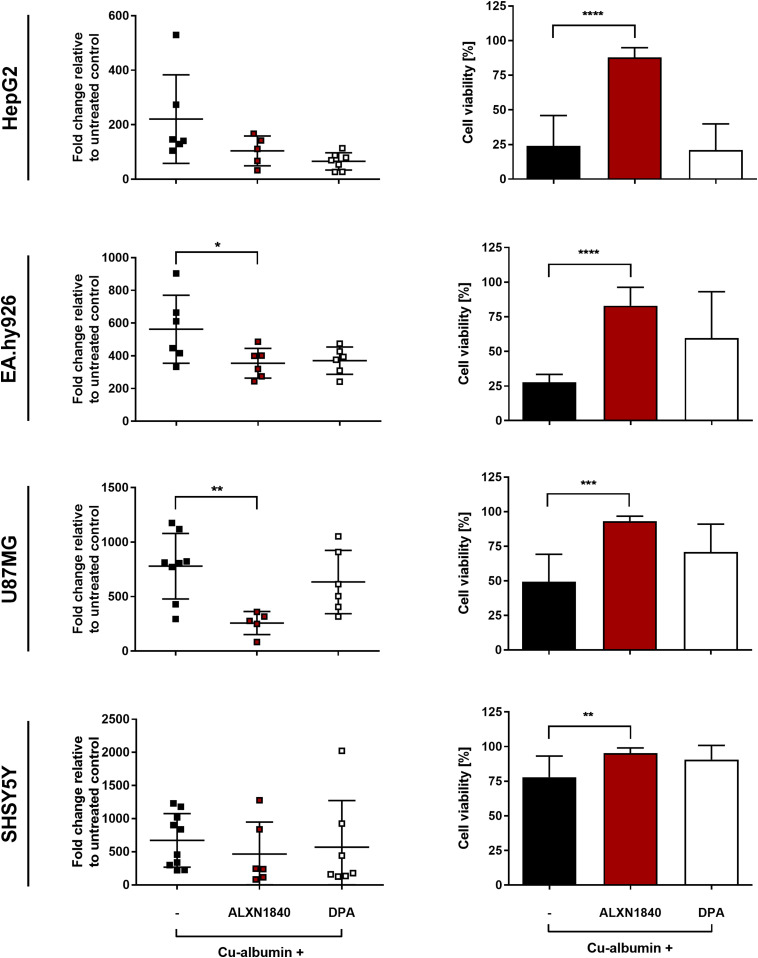
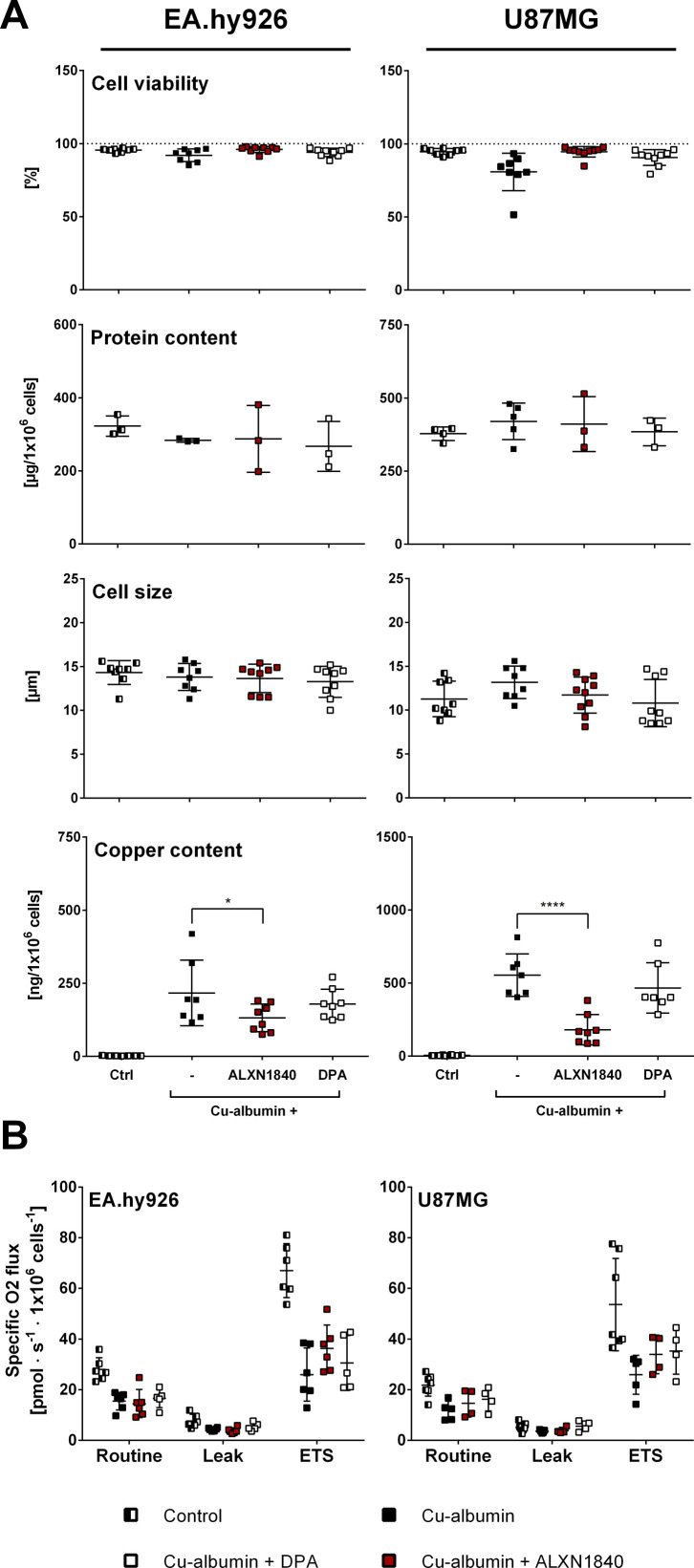
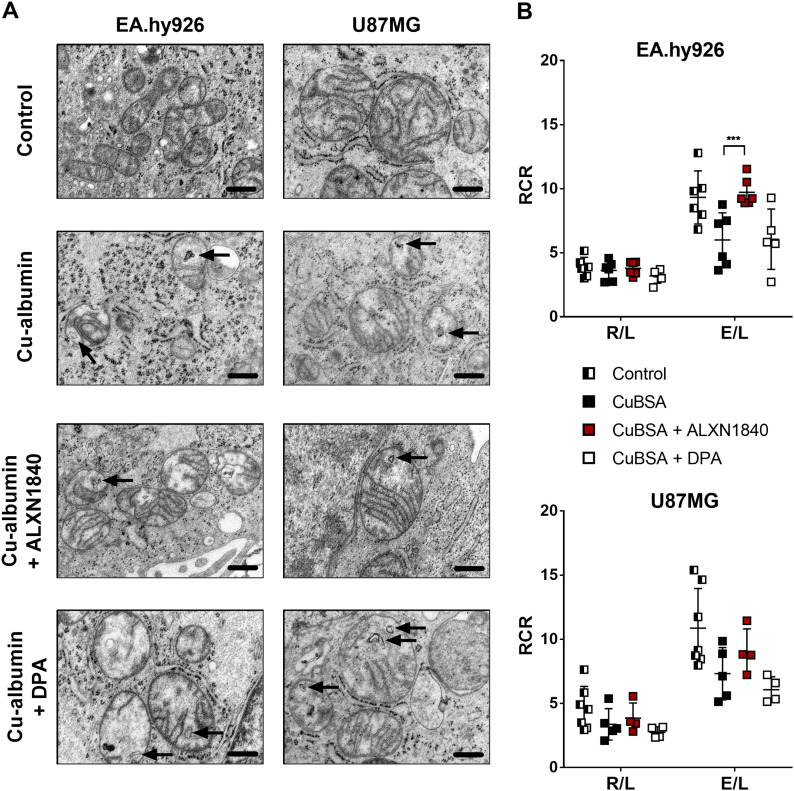
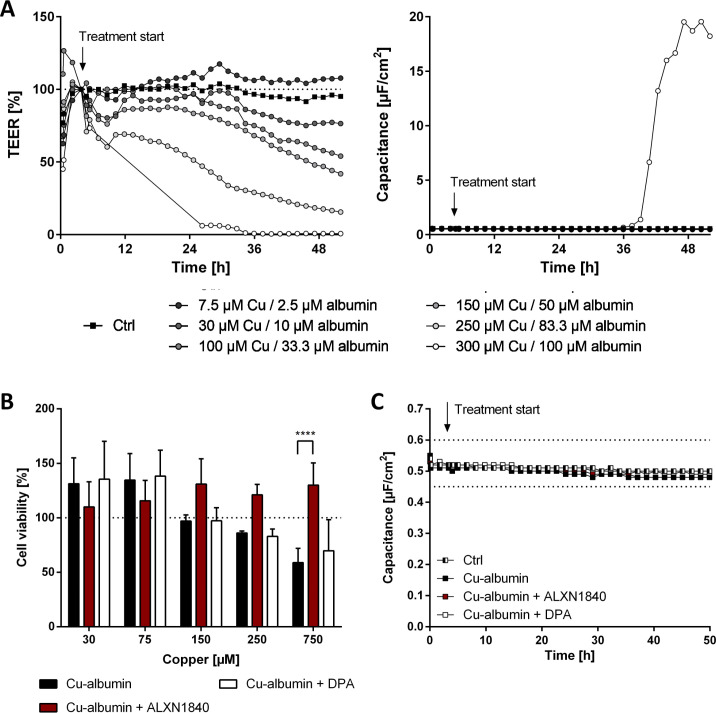
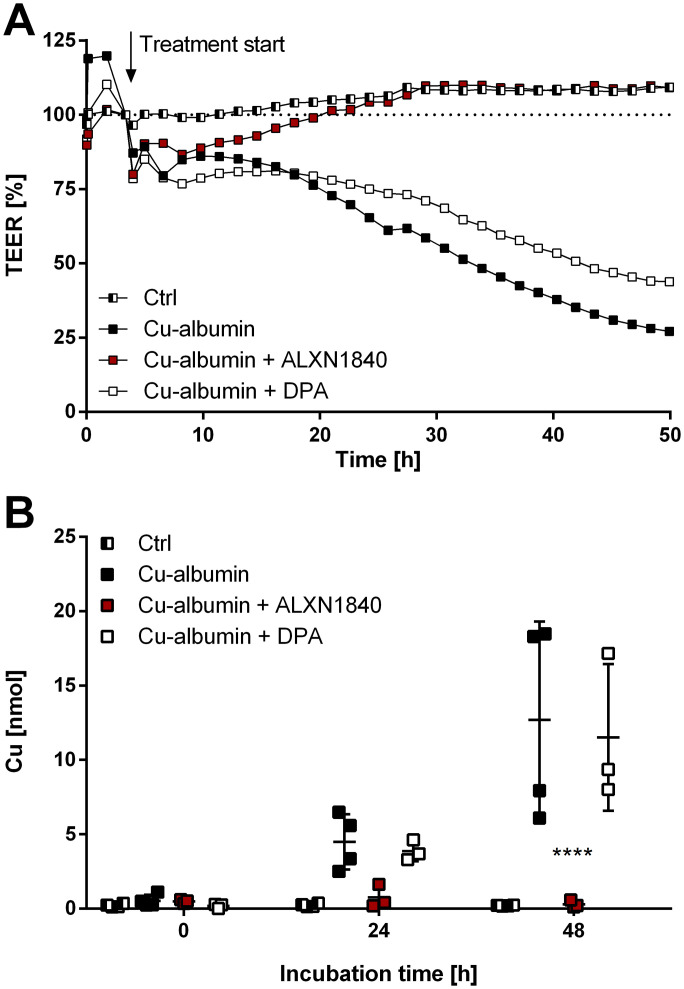
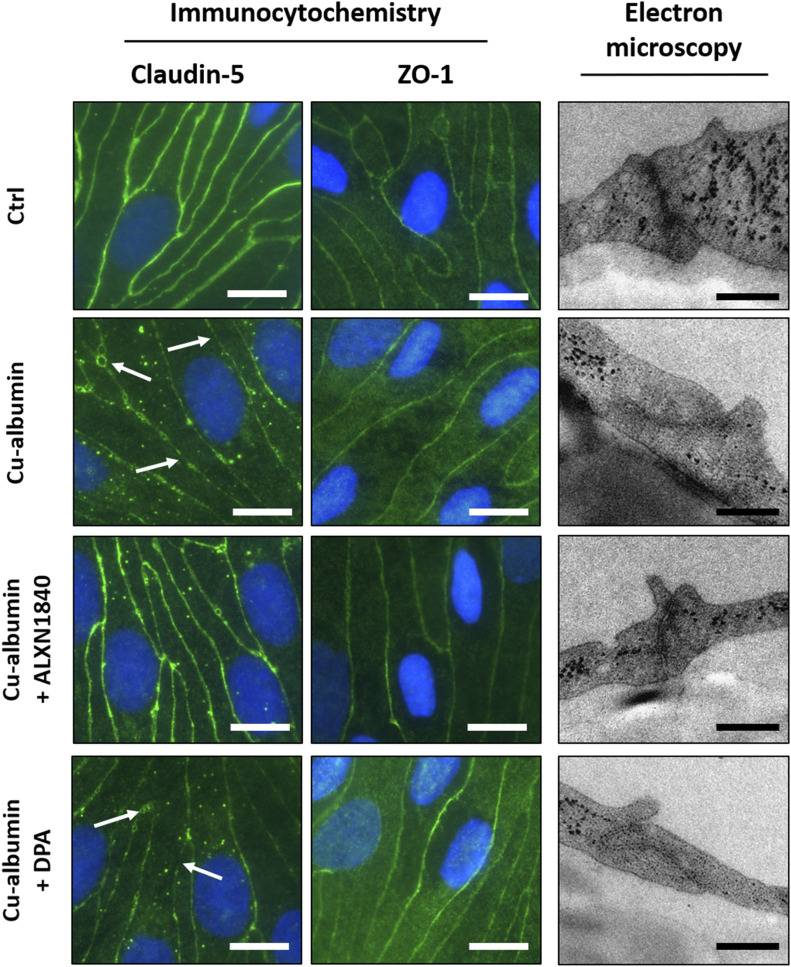
In Wilson disease, excessive copper accumulates in patients' livers and may, upon serum leakage, severely affect the brain according to current viewpoints. Present remedies aim at avoiding copper toxicity by chelation, for example, by D-penicillamine (DPA) or bis-choline tetrathiomolybdate (ALXN1840), the latter with a very high copper affinity. Hence, ALXN1840 may potentially avoid neurological deterioration that frequently occurs upon DPA treatment. As the etiology of such worsening is unclear, we reasoned that copper loosely bound to albumin, that is, mimicking a potential liver copper leakage into blood, may damage cells that constitute the blood-brain barrier, which was found to be the case in an in vitro model using primary porcine brain capillary endothelial cells. Such blood-brain barrier damage was avoided by ALXN1840, plausibly due to firm protein embedding of the chelator bound copper, but not by DPA. Mitochondrial protection was observed, a prerequisite for blood-brain barrier integrity. Thus, high-affinity copper chelators may minimize such deterioration in the treatment of neurologic Wilson disease.
© 2021 Borchard et al.
Conflict of interest statement
AH Coles and R Viana are employees and shareholders of Alexion AstraZeneca Rare Disease. T Plitz was an employee of Wilson Therapeutics.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous