

Retroviral integrase: Structure, mechanism, and inhibition
- PMID: 34861940
- PMCID: PMC8732146
- DOI: 10.1016/bs.enz.2021.06.007
Retroviral integrase: Structure, mechanism, and inhibition
Abstract
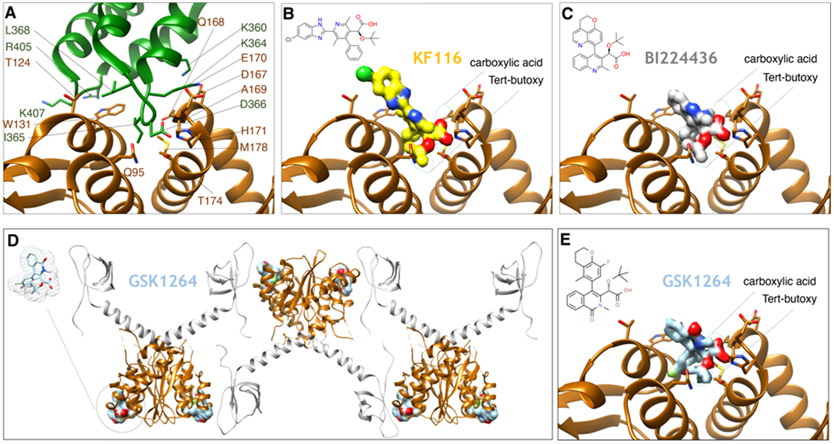
The retroviral protein Integrase (IN) catalyzes concerted integration of viral DNA into host chromatin to establish a permanent infection in the target cell. We learned a great deal about the mechanism of catalytic integration through structure/function studies over the previous four decades of IN research. As one of three essential retroviral enzymes, IN has also been targeted by antiretroviral drugs to treat HIV-infected individuals. Inhibitors blocking the catalytic integration reaction are now state-of-the-art drugs within the antiretroviral therapy toolkit. HIV-1 IN also performs intriguing non-catalytic functions that are relevant to the late stages of the viral replication cycle, yet this aspect remains poorly understood. There are also novel allosteric inhibitors targeting non-enzymatic functions of IN that induce a block in the late stages of the viral replication cycle. In this chapter, we will discuss the function, structure, and inhibition of retroviral IN proteins, highlighting remaining challenges and outstanding questions.
Keywords: Antiretroviral therapy; Drug design; Enzyme; Hydrolase; Retrovirus.
Copyright © 2021 Elsevier Inc. All rights reserved.
Figures
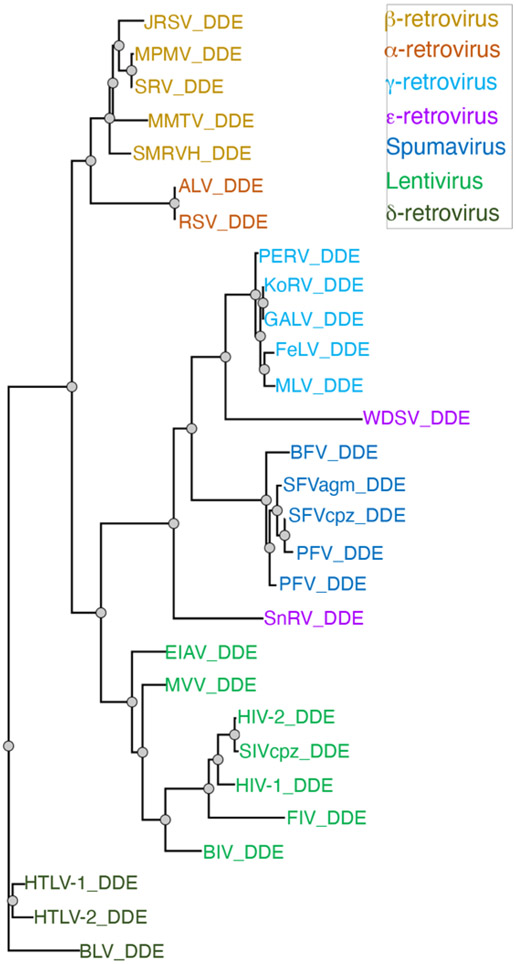
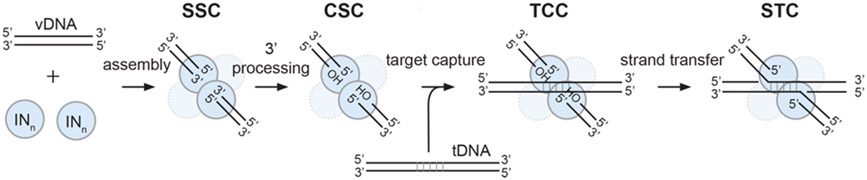
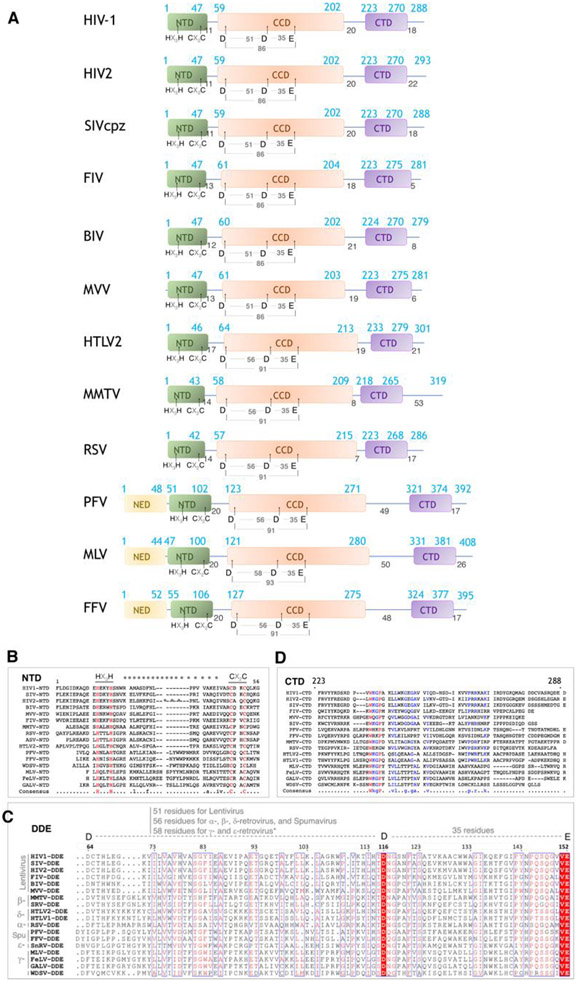
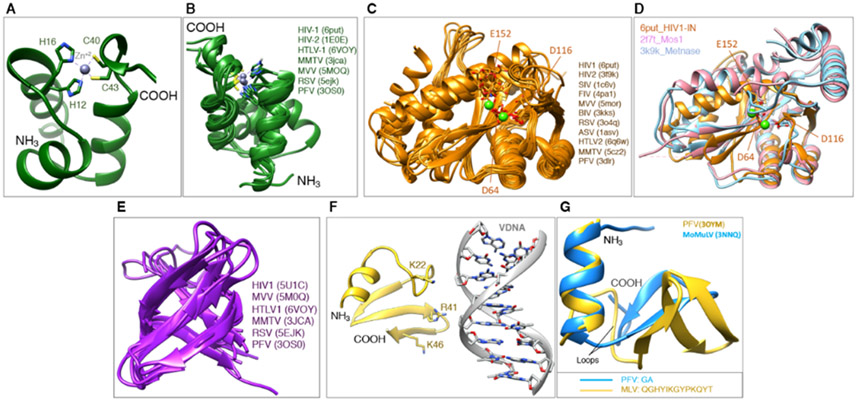
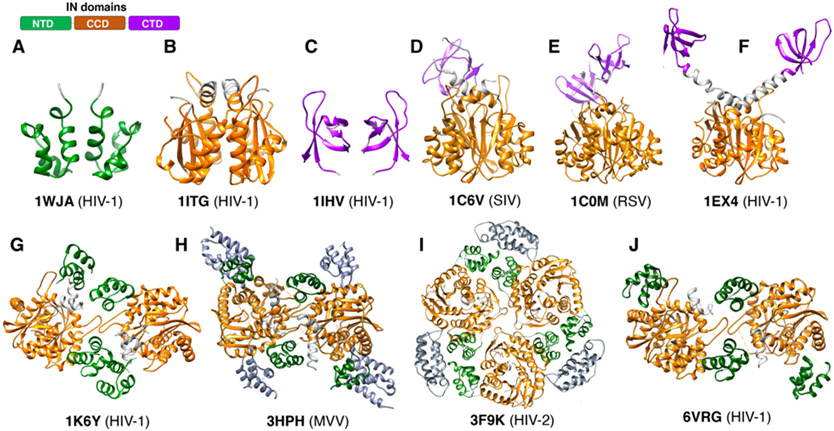
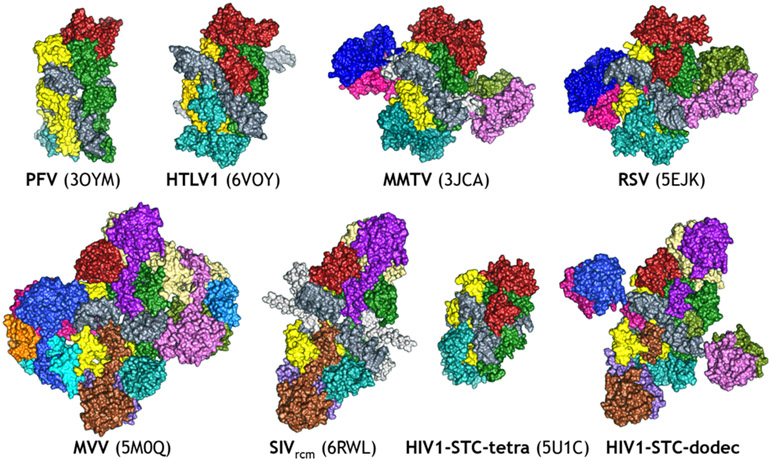
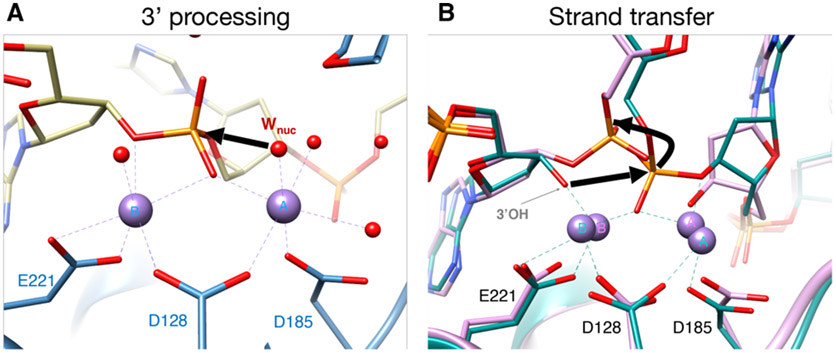
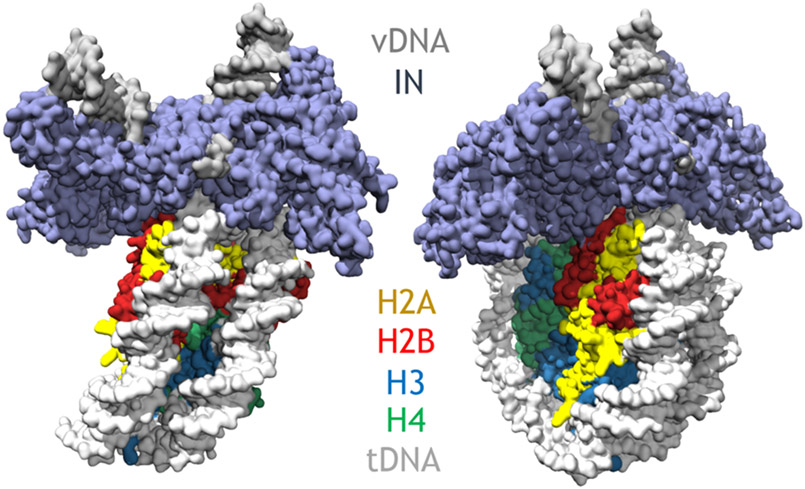
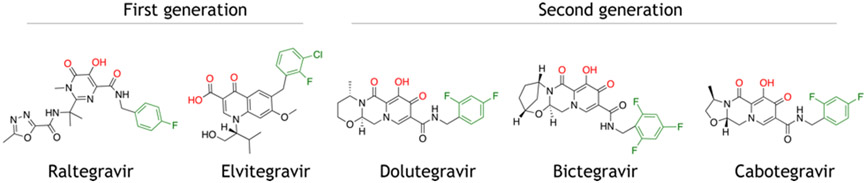
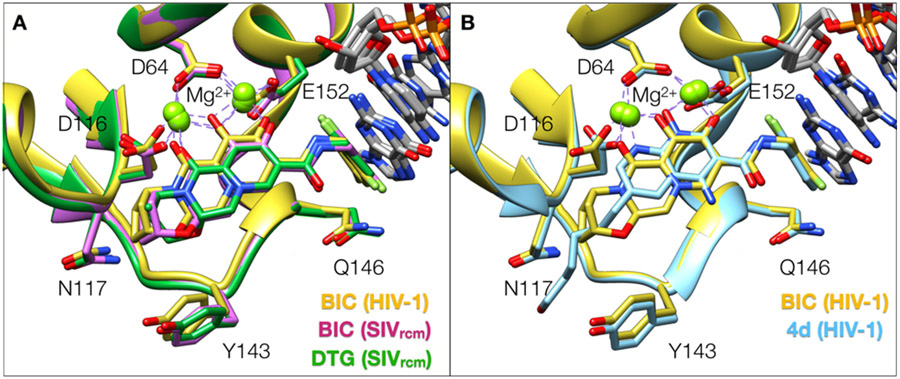
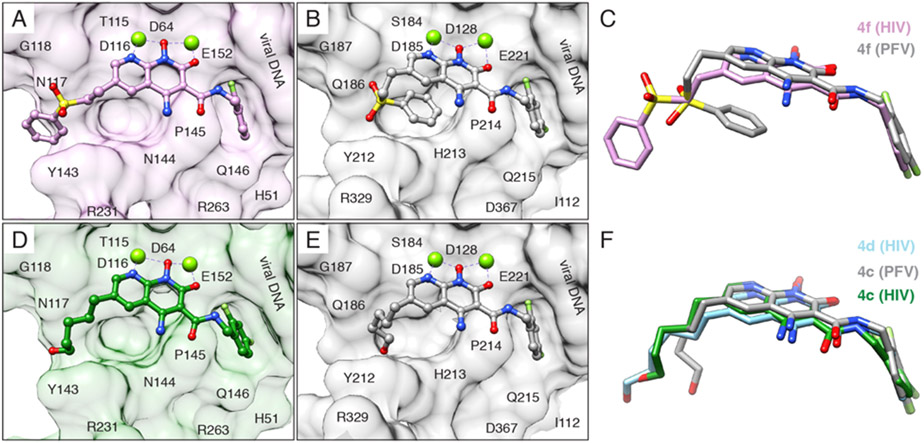
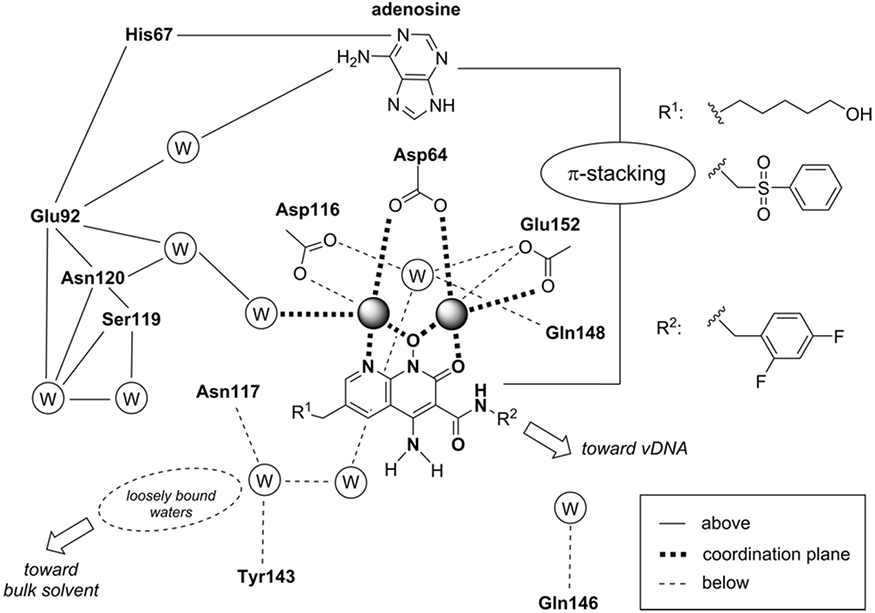
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials