

Targeting cancer metabolism in the era of precision oncology
- PMID: 34862480
- PMCID: PMC8641543
- DOI: 10.1038/s41573-021-00339-6
Targeting cancer metabolism in the era of precision oncology
Abstract
One hundred years have passed since Warburg discovered alterations in cancer metabolism, more than 70 years since Sidney Farber introduced anti-folates that transformed the treatment of childhood leukaemia, and 20 years since metabolism was linked to oncogenes. However, progress in targeting cancer metabolism therapeutically in the past decade has been limited. Only a few metabolism-based drugs for cancer have been successfully developed, some of which are in - or en route to - clinical trials. Strategies for targeting the intrinsic metabolism of cancer cells often did not account for the metabolism of non-cancer stromal and immune cells, which have pivotal roles in tumour progression and maintenance. By considering immune cell metabolism and the clinical manifestations of inborn errors of metabolism, it may be possible to isolate undesirable off-tumour, on-target effects of metabolic drugs during their development. Hence, the conceptual framework for drug design must consider the metabolic vulnerabilities of non-cancer cells in the tumour immune microenvironment, as well as those of cancer cells. In this Review, we cover the recent developments, notable milestones and setbacks in targeting cancer metabolism, and discuss the way forward for the field.
© 2021. Springer Nature Limited.
Conflict of interest statement
C.V.D. is a consultant of the Barer Institute, Inc. and member of the board of directors of Rafael Pharmaceuticals, Inc. J.S. receives sponsored research support from the Barer Institute, Inc. Z.E.S. is an employee of the Barer Institute. Z.T.S. declares no competing interests.
Figures
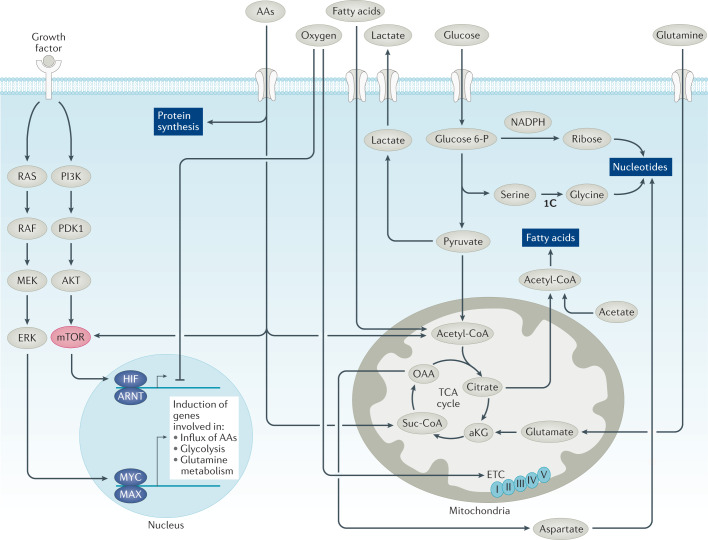
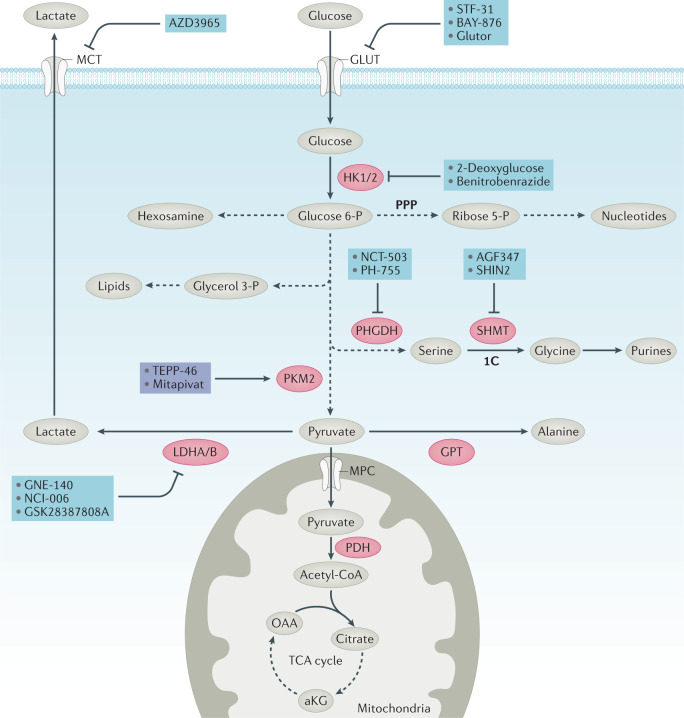
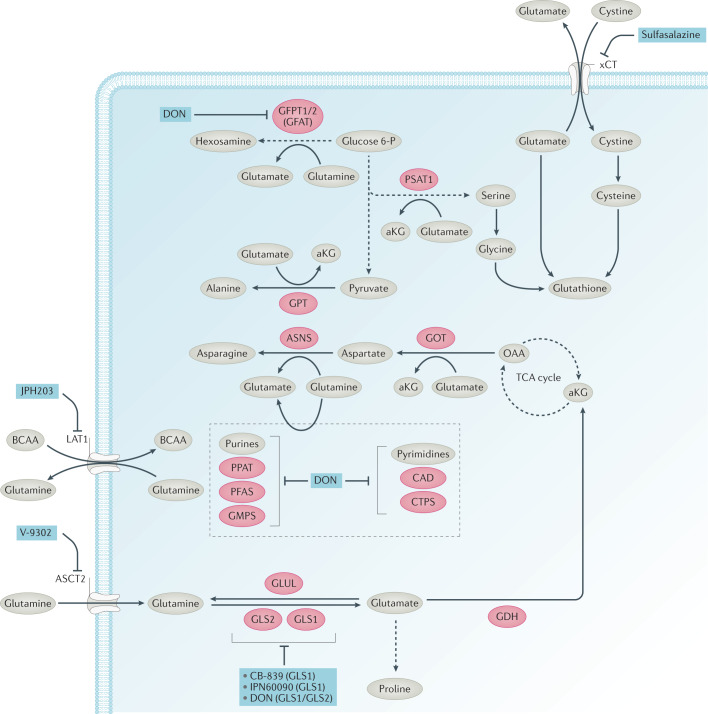
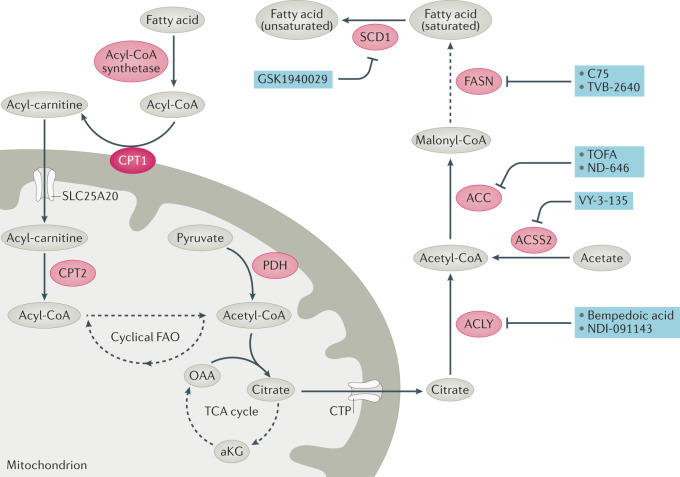
References
-
- Farber S, Diamond LK. Temporary remissions in acute leukemia in children produced by folic acid antagonist, 4-aminopteroyl-glutamic acid. N. Engl. J. Med. 1948;238:787–793. - PubMed
-
- Warburg O. On respiratory impairment in cancer cells. Science. 1956;124:269–270. - PubMed
-
- Landau BR, Laszlo J, Stengle J, Burk D. Certain metabolic and pharmacologic effects in cancer patients given infusions of 2-deoxy-d-glucose. J. Natl Cancer Inst. 1958;21:485–494. - PubMed
-
- Miller DR. A tribute to Sidney Farber — the father of modern chemotherapy. Br. J. Haematol. 2006;134:20–26. - PubMed
-
- Dang CV, Semenza GL. Oncogenic alterations of metabolism. Trends Biochem. Sci. 1999;24:68–72. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
