

A Pulmonary Vascular Model From Endothelialized Whole Organ Scaffolds
- PMID: 34869270
- PMCID: PMC8640093
- DOI: 10.3389/fbioe.2021.760309
A Pulmonary Vascular Model From Endothelialized Whole Organ Scaffolds
Abstract
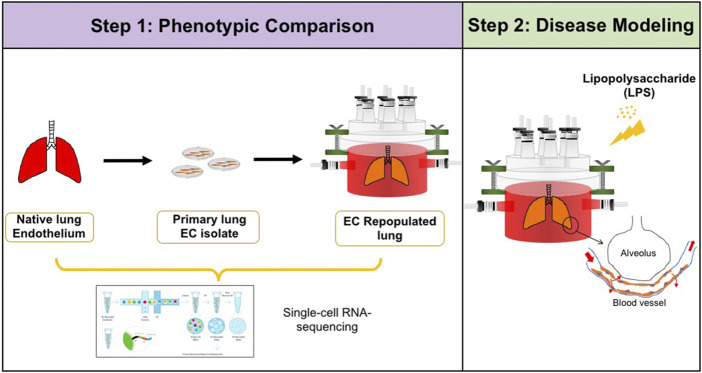
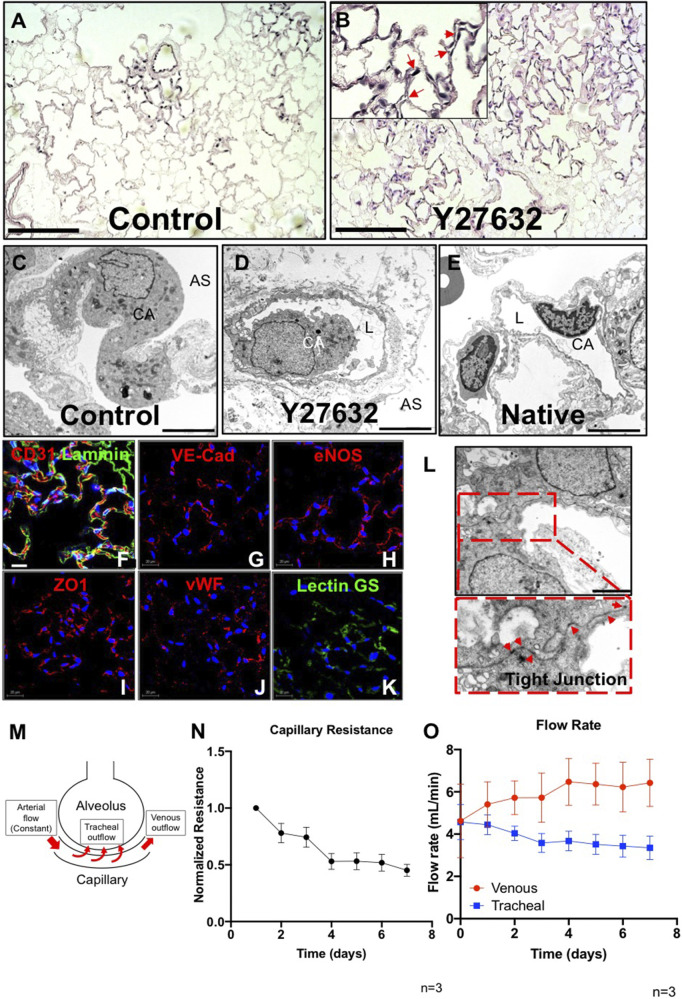
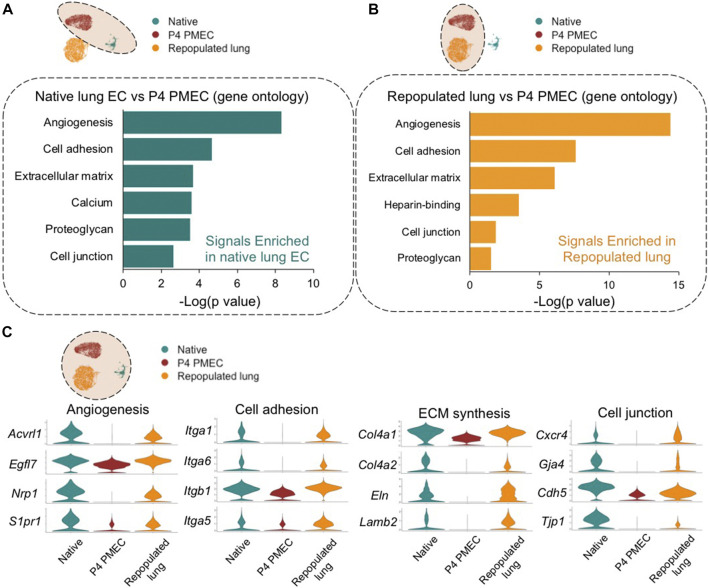
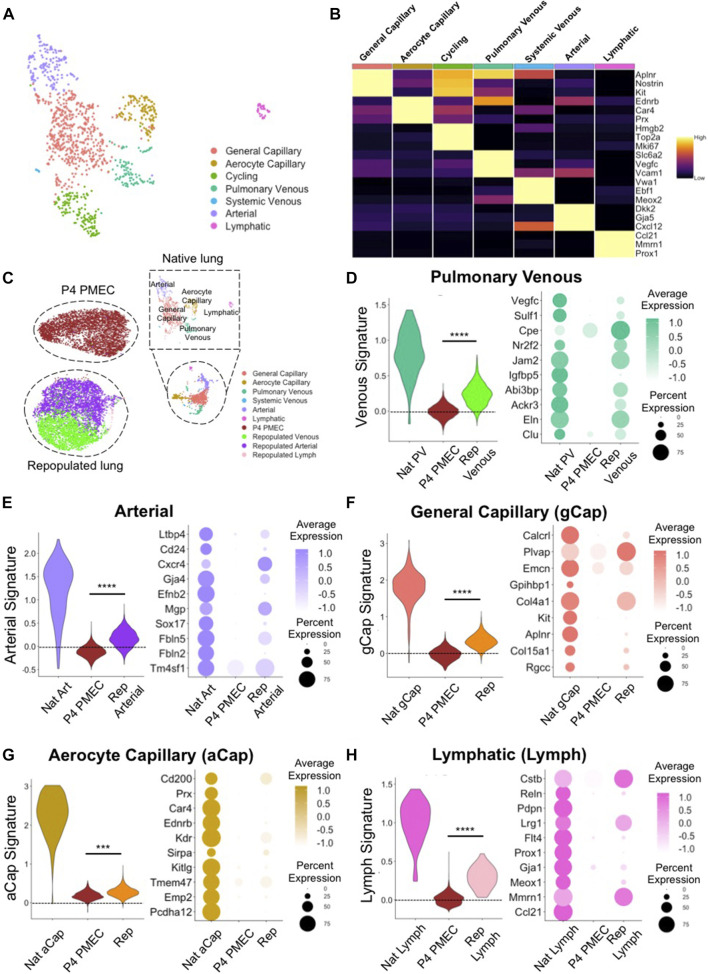
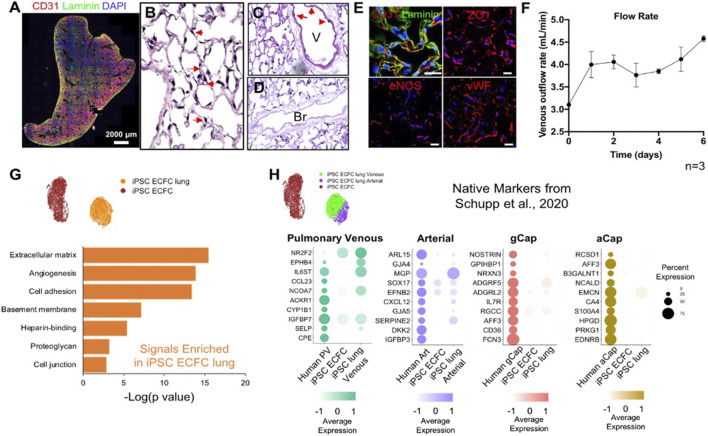
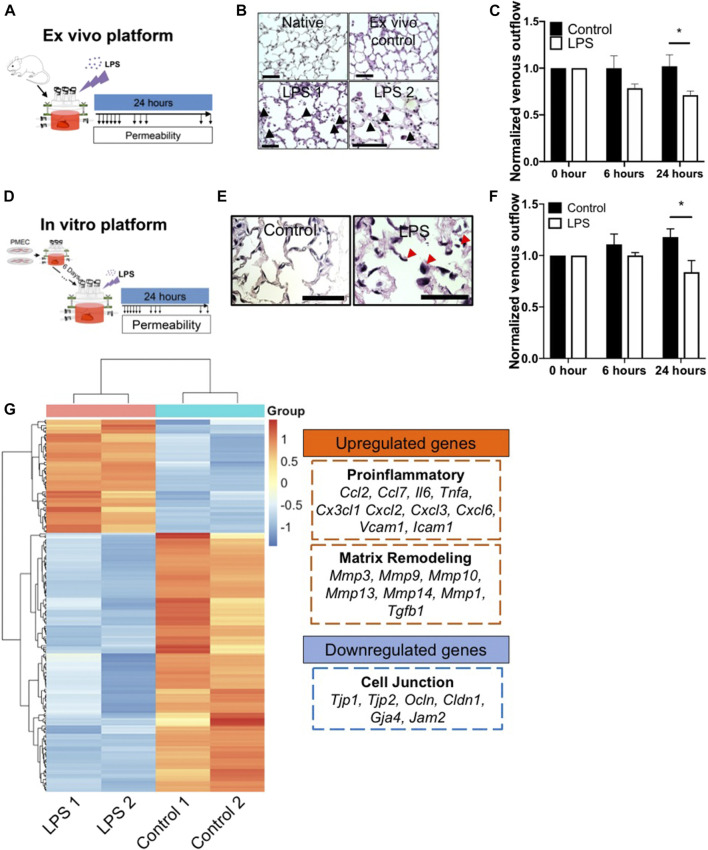
The development of an in vitro system for the study of lung vascular disease is critical to understanding human pathologies. Conventional culture systems fail to fully recapitulate native microenvironmental conditions and are typically limited in their ability to represent human pathophysiology for the study of disease and drug mechanisms. Whole organ decellularization provides a means to developing a construct that recapitulates structural, mechanical, and biological features of a complete vascular structure. Here, we developed a culture protocol to improve endothelial cell coverage in whole lung scaffolds and used single-cell RNA-sequencing analysis to explore the impact of decellularized whole lung scaffolds on endothelial phenotypes and functions in a biomimetic bioreactor system. Intriguingly, we found that the phenotype and functional signals of primary pulmonary microvascular revert back-at least partially-toward native lung endothelium. Additionally, human induced pluripotent stem cell-derived endothelium cultured in decellularized lung systems start to gain various native human endothelial phenotypes. Vascular barrier function was partially restored, while small capillaries remained patent in endothelial cell-repopulated lungs. To evaluate the ability of the engineered endothelium to modulate permeability in response to exogenous stimuli, lipopolysaccharide (LPS) was introduced into repopulated lungs to simulate acute lung injury. After LPS treatment, proinflammatory signals were significantly increased and the vascular barrier was impaired. Taken together, these results demonstrate a novel platform that recapitulates some pulmonary microvascular functions and phenotypes at a whole organ level. This development may help pave the way for using the whole organ engineering approach to model vascular diseases.
Keywords: endothelium; in vitro disease modeling; pulmonary vasculature; single-cell RNA-sequencing; whole lung tissue engineering.
Copyright © 2021 Yuan, Leiby, Greaney, Raredon, Qian, Schupp, Engler, Baevova, Adams, Kural, Wang, Obata, Yoder, Kaminski and Niklason.
Conflict of interest statement
LEN is a founder and shareholder in Humacyte, Inc. The remaining authors declare that the research was conducted in the Q14 absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases