

Immunodeficiency Accelerates Vitamin A Deficiency
- PMID: 34870072
- PMCID: PMC8634461
- DOI: 10.1093/cdn/nzab129
Immunodeficiency Accelerates Vitamin A Deficiency
Abstract
Background: Vitamin A deficiency increases susceptibility to infection caused by impaired immune function.
Objectives: We investigated whether immunodeficiency could facilitate the development of vitamin A deficiency.
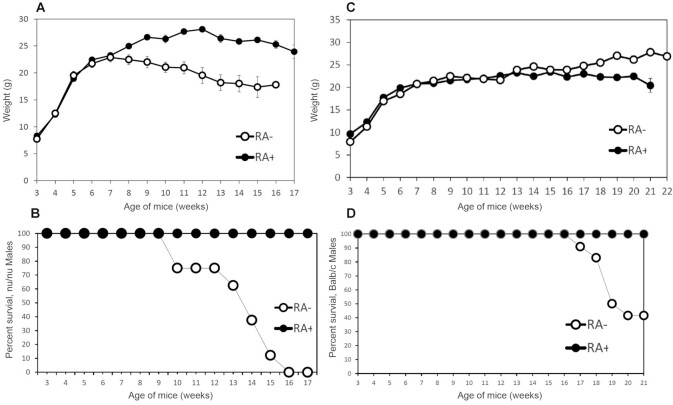
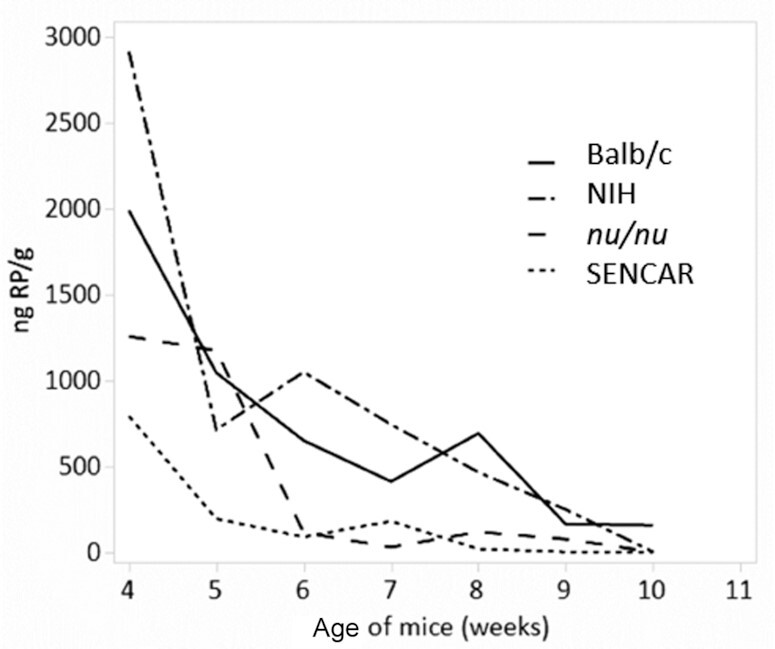
Methods: Vitamin A deficiency was followed in 2 mouse models of immunodeficiency: the athymic nude mouse (nu/nu) and the humoral immunodeficient SENCAR (SENsitive to CARcinogenesis) mouse. Vitamin A deficiency was also monitored in outbred Balb/c and in NIH mice. The monitoring of vitamin A deficiency was done after feeding the mice and their mothers a semisynthetic, vitamin A-deficient diet from birth of the experimental mice. These mice were weaned onto the same deficient diet at 3-4 wk of age, while control groups were fed the same diet containing 3 μg retinoic acid per gram of diet.
Results: The immunodeficient nu/nu and SENCAR mice developed vitamin A deficiency earlier than either the heterozygous nu/+ controls or the Balb/c and NIH strains. In female mice, symptoms included depletion of liver retinol and retinyl palmitate, squamous metaplasia of the uterus, and death. Male mice lost weight more frequently and sooner than female mice, in which mortality generally occurred in the absence of loss of body weight. Pairwise comparisons using Tukey's honest significant difference test of the nu/nu and SENCAR mice versus the Balb/c and NIH mice showed a faster loss of retinol and retinyl palmitate in all pairs (P ≤ 0.0001) except for retinol when comparing nu/nu and NIH strains (P = 0.3383).
Conclusions: Our findings are consistent with an increased usage of liver retinol and retinyl palmitate in the immunocompromised nu/nu and in the immunodeficient SENCAR mice and suggest that compensatory mechanisms dependent on vitamin A utilization are called upon to rescue immunodeficiency both in the T-cell-deficient phenotype of the nu/nu mice and in the humoral immunodeficient SENCAR mice.
Keywords: SENCAR (SENsitive to CARcinogenesis) mice; athymic mice; immunodeficiency; retinol; retinyl palmitate; vitamin A deficiency.
© The Author(s) 2021. Published by Oxford University Press on behalf of the American Society for Nutrition.
Figures









References
LinkOut - more resources
Full Text Sources
Research Materials
