

Proteolytic activation of the epithelial sodium channel (ENaC) by factor VII activating protease (FSAP) and its relevance for sodium retention in nephrotic mice
- PMID: 34870751
- PMCID: PMC8766372
- DOI: 10.1007/s00424-021-02639-7
Proteolytic activation of the epithelial sodium channel (ENaC) by factor VII activating protease (FSAP) and its relevance for sodium retention in nephrotic mice
Abstract
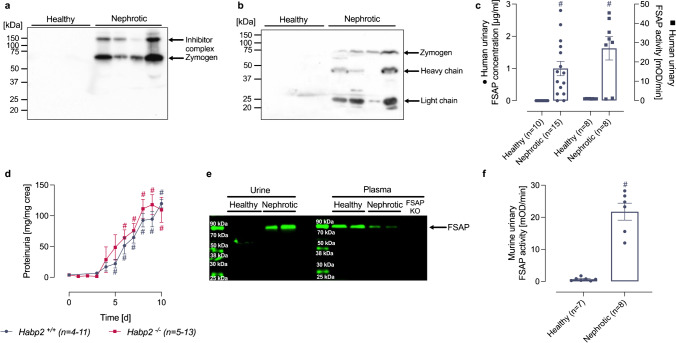
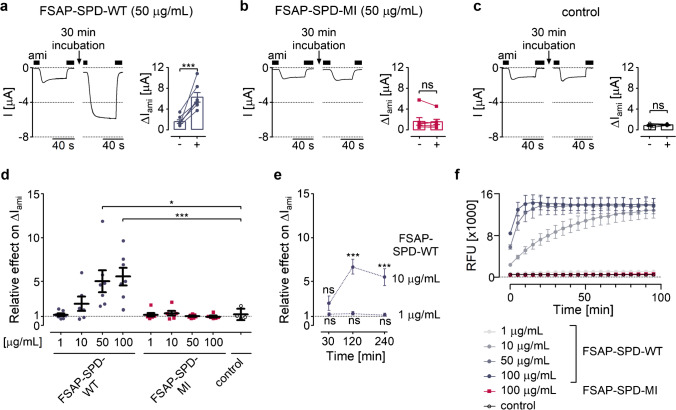
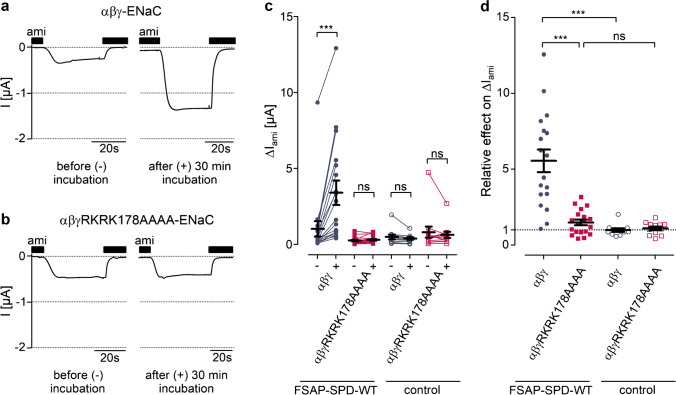
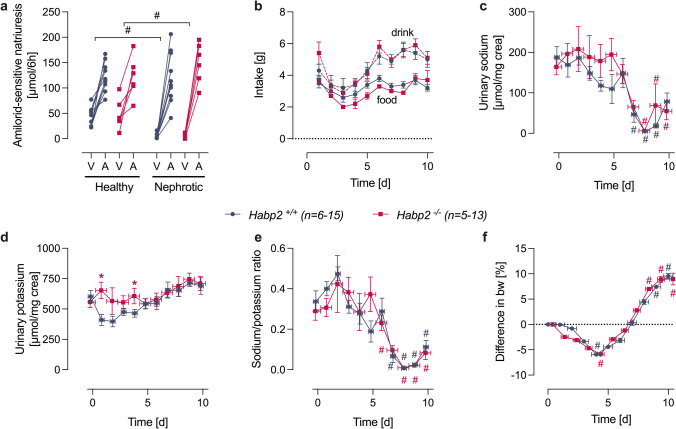
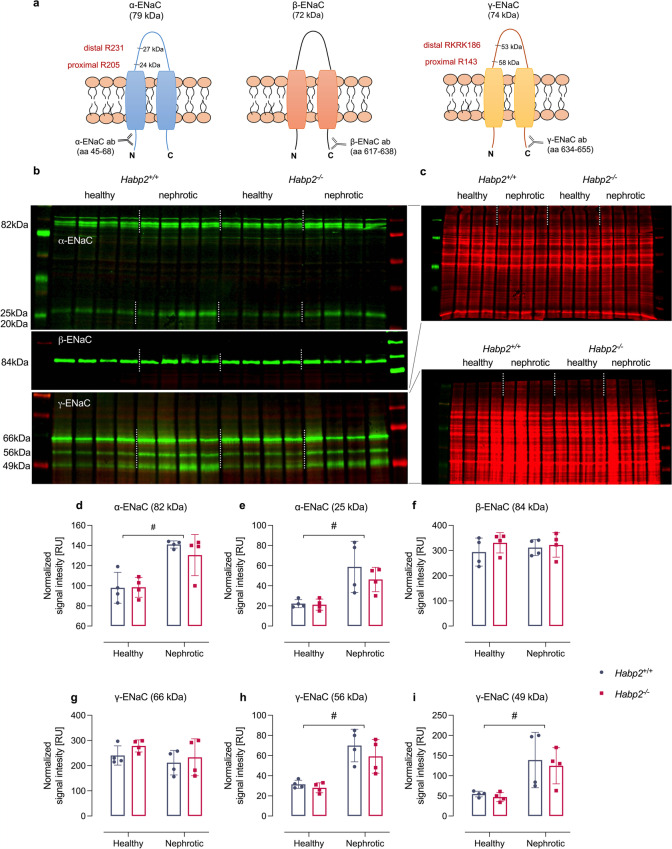
Proteolytic activation of the epithelial sodium channel (ENaC) by aberrantly filtered serine proteases is thought to contribute to renal sodium retention in nephrotic syndrome. However, the identity of the responsible proteases remains elusive. This study evaluated factor VII activating protease (FSAP) as a candidate in this context. We analyzed FSAP in the urine of patients with nephrotic syndrome and nephrotic mice and investigated its ability to activate human ENaC expressed in Xenopus laevis oocytes. Moreover, we studied sodium retention in FSAP-deficient mice (Habp2-/-) with experimental nephrotic syndrome induced by doxorubicin. In urine samples from nephrotic humans, high concentrations of FSAP were detected both as zymogen and in its active state. Recombinant serine protease domain of FSAP stimulated ENaC-mediated whole-cell currents in a time- and concentration-dependent manner. Mutating the putative prostasin cleavage site in γ-ENaC (γRKRK178AAAA) prevented channel stimulation by the serine protease domain of FSAP. In a mouse model for nephrotic syndrome, active FSAP was present in nephrotic urine of Habp2+/+ but not of Habp2-/- mice. However, Habp2-/- mice were not protected from sodium retention compared to nephrotic Habp2+/+ mice. Western blot analysis revealed that in nephrotic Habp2-/- mice, proteolytic cleavage of α- and γ-ENaC was similar to that in nephrotic Habp2+/+ animals. In conclusion, active FSAP is excreted in the urine of nephrotic patients and mice and activates ENaC in vitro involving the putative prostasin cleavage site of γ-ENaC. However, endogenous FSAP is not essential for sodium retention in nephrotic mice.
Keywords: Epithelial sodium channel (ENaC); FSAP-HABP2; Factor VII activating protease; Nephrotic syndrome; Serine protease.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Bohnert BN, Daiminger S, Worn M, Sure F, Staudner T, Ilyaskin AV, Batbouta F, Janessa A, Schneider JC, Essigke D, Kanse S, Haerteis S, Korbmacher C, Artunc F. Urokinase-type plasminogen activator (uPA) is not essential for epithelial sodium channel (ENaC)-mediated sodium retention in experimental nephrotic syndrome. Acta Physiol (Oxf) 2019;227:e13286. doi: 10.1111/apha.13286. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
