

New Delhi Metallo-Beta-Lactamase Facilitates the Emergence of Cefiderocol Resistance in Enterobacter cloacae
- PMID: 34871093
- PMCID: PMC8846454
- DOI: 10.1128/AAC.02011-21
New Delhi Metallo-Beta-Lactamase Facilitates the Emergence of Cefiderocol Resistance in Enterobacter cloacae
Abstract
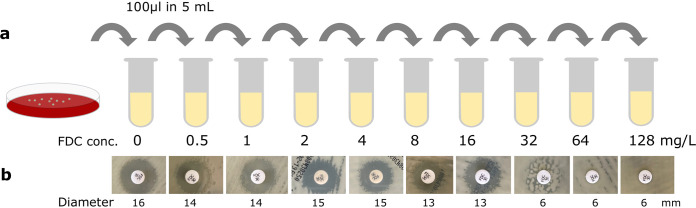
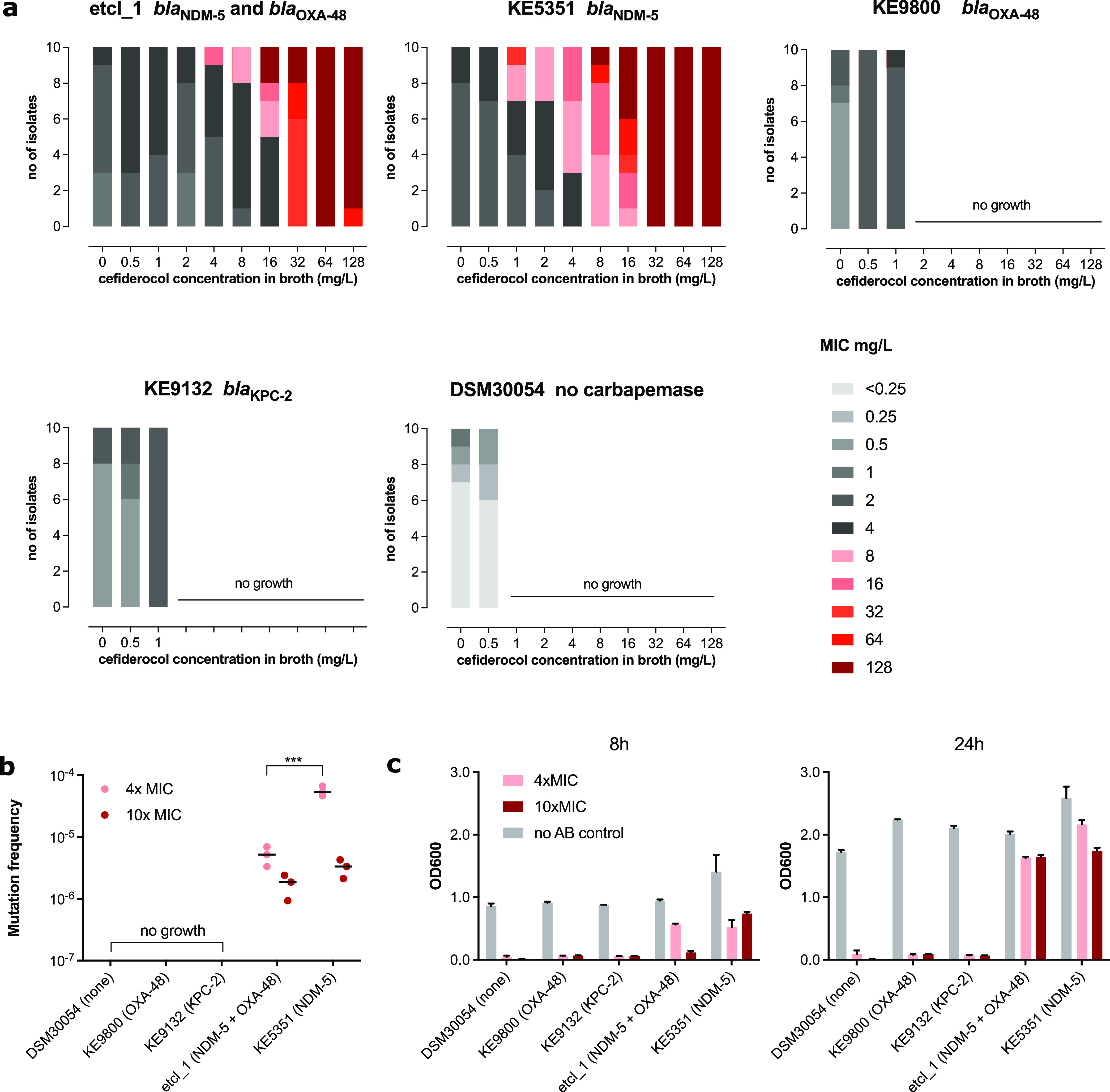
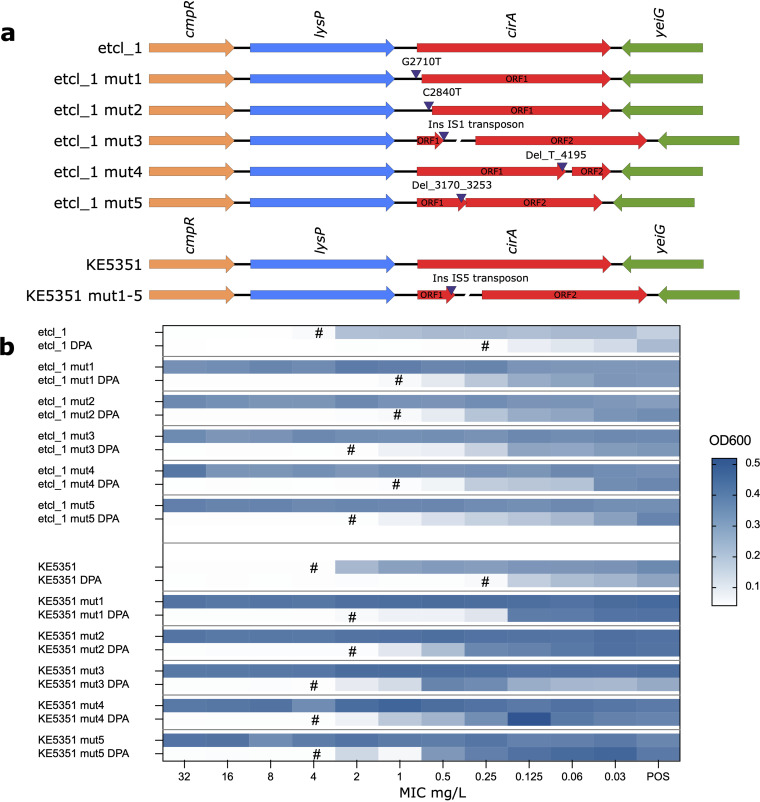
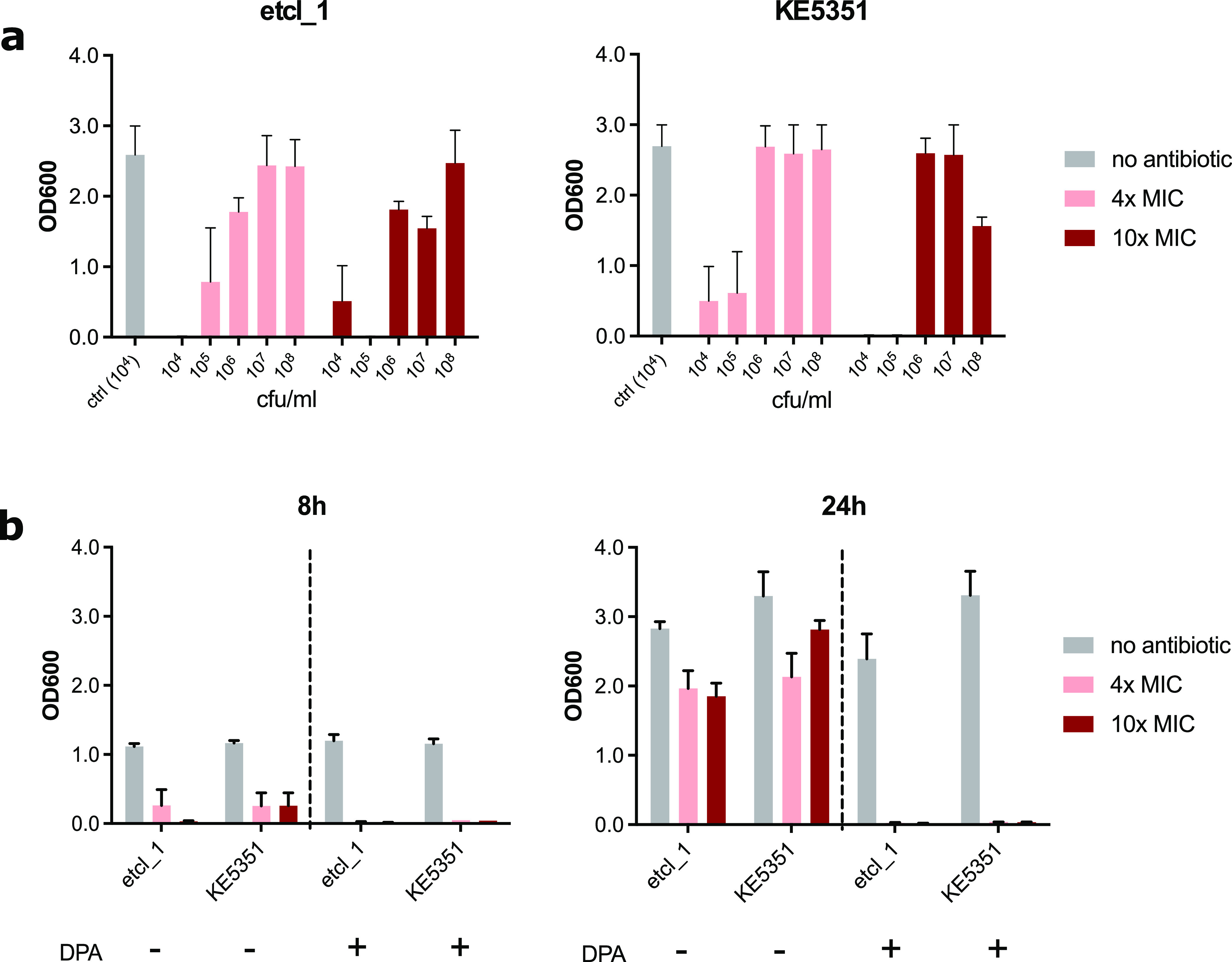
Cefiderocol is a promising novel siderophore cephalosporin for the treatment of multidrug-resistant Gram-negative bacilli and with stability against degradation by metallo-β-lactamases. Nonetheless, the emergence of cefiderocol in metallo-β-lactamase-producing Enterobacterales during therapy has been reported on more than one occasion. To understand the underlying mechanisms and factors facilitating the resistance development, we conducted an in vitro evolution experiment using clinical E. cloacae isolates via serial passaging under cefiderocol pressure. In this study, we showed that the presence of the New Delhi metallo-β-lactamase (NDM) facilitates the emergence of resistance via nonsynonymous mutations of the CirA catecholate siderophore receptor. Inhibition of metallo-β-lactamase activity using dipicolinic acid prevented the emergence of cefiderocol-resistant mutants successfully. This finding implies that caution should be taken when using cefiderocol for the treatment of infections caused by metallo-β-lactamase-producing bacteria.
Keywords: Enterobacter cloacae; Enterobacterales; New Delhi metallo-β-lactamase; antibiotic resistance; carbapenem resistance; cefiderocol; cefiderocol resistance; drug resistance mechanisms.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Cassini A, Högberg LD, Plachouras D, Quattrocchi A, Hoxha A, Simonsen GS, Colomb-Cotinat M, Kretzschmar ME, Devleesschauwer B, Cecchini M, Ouakrim DA, Oliveira TC, Struelens MJ, Suetens C, Monnet DL, Strauss R, Mertens K, Struyf T, Catry B, Latour K, Ivanov IN, Dobreva EG, Tambic Andraševic A, Soprek S, Budimir A, Paphitou N, Žemlicková H, Schytte Olsen S, Wolff Sönksen U, Märtin P, Ivanova M, Lyytikäinen O, Jalava J, Coignard B, Eckmanns T, Abu Sin M, Haller S, Daikos GL, Gikas A, Tsiodras S, Kontopidou F, Tóth Á, Hajdu Á, Guólaugsson Ó, Kristinsson KG, Murchan S, Burns K, Pezzotti P, Gagliotti C, Dumpis U, Burden of AMRCG, et al.. 2019. Attributable deaths and disability-adjusted life-years caused by infections with antibiotic-resistant bacteria in the EU and the European Economic Area in 2015: a population-level modelling analysis. Lancet Infect Dis 19:56–66. 10.1016/S1473-3099(18)30605-4. - DOI - PMC - PubMed
-
- Tacconelli E, Carrara E, Savoldi A, Harbarth S, Mendelson M, Monnet DL, Pulcini C, Kahlmeter G, Kluytmans J, Carmeli Y, Ouellette M, Outterson K, Patel J, Cavaleri M, Cox EM, Houchens CR, Grayson ML, Hansen P, Singh N, Theuretzbacher U, Magrini N, Group WHOPPLW. 2018. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis 18:318–327. 10.1016/S1473-3099(17)30753-3. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
