

Stem-like intestinal Th17 cells give rise to pathogenic effector T cells during autoimmunity
- PMID: 34875227
- PMCID: PMC8900676
- DOI: 10.1016/j.cell.2021.11.018
Stem-like intestinal Th17 cells give rise to pathogenic effector T cells during autoimmunity
Abstract
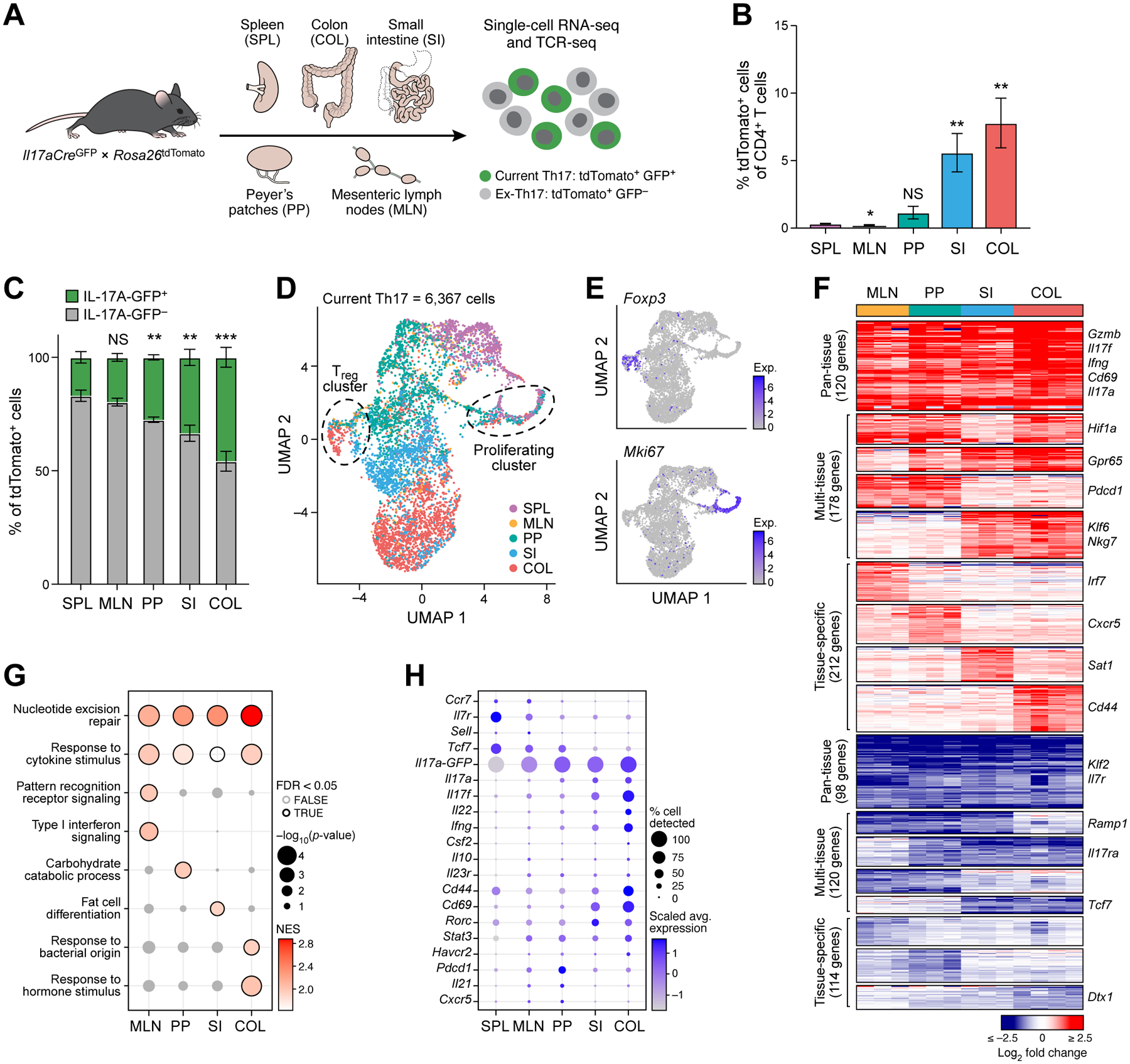
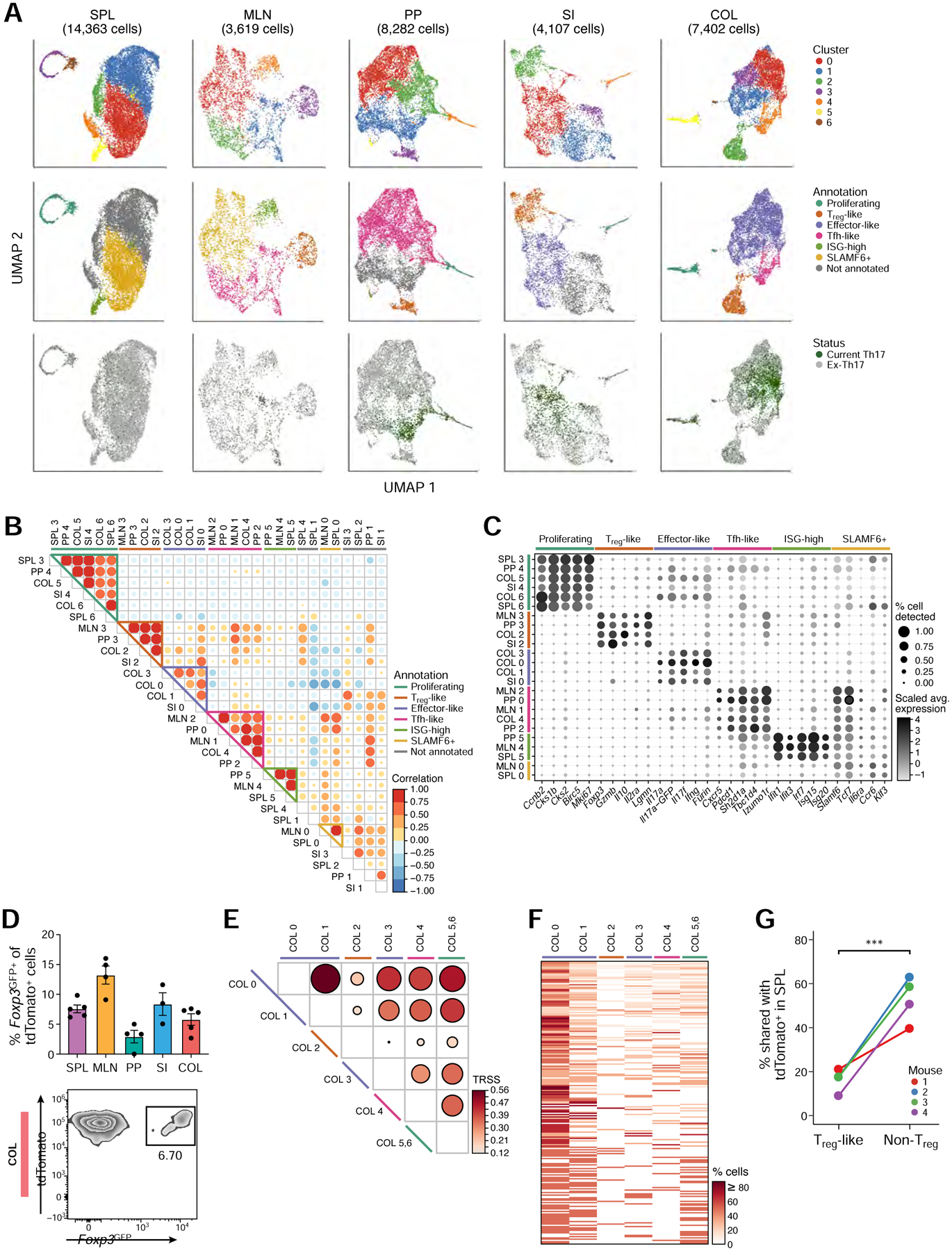
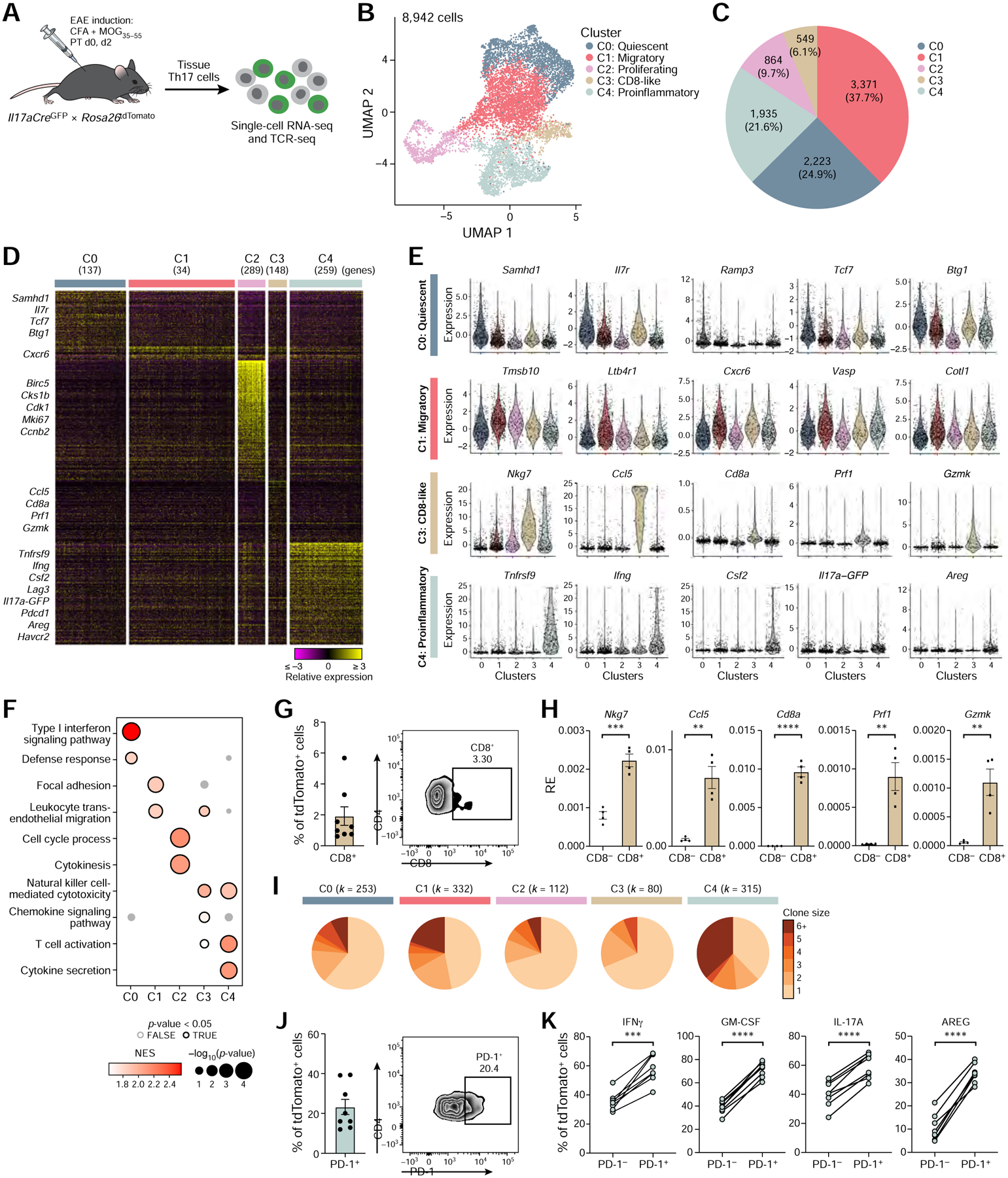
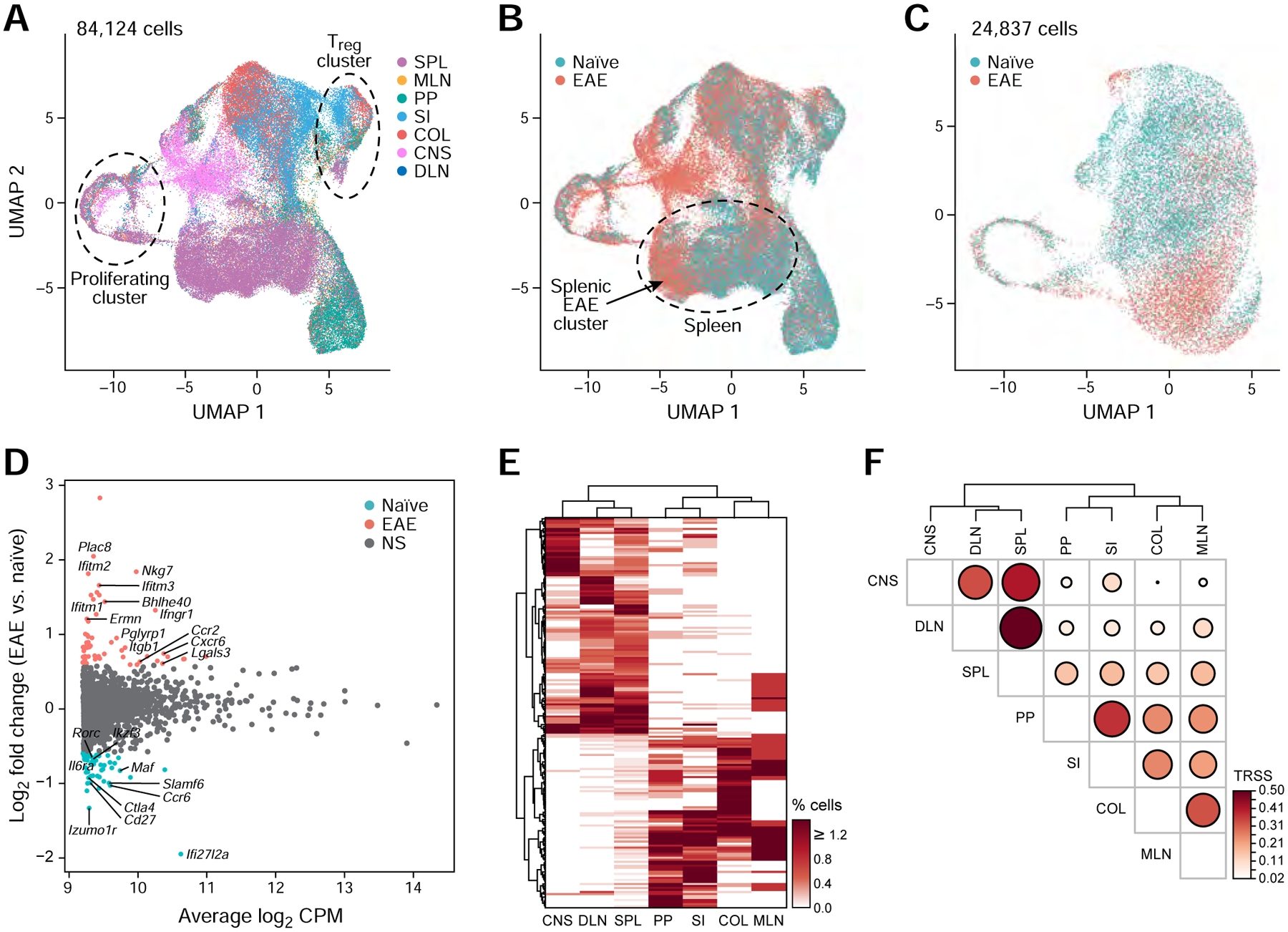
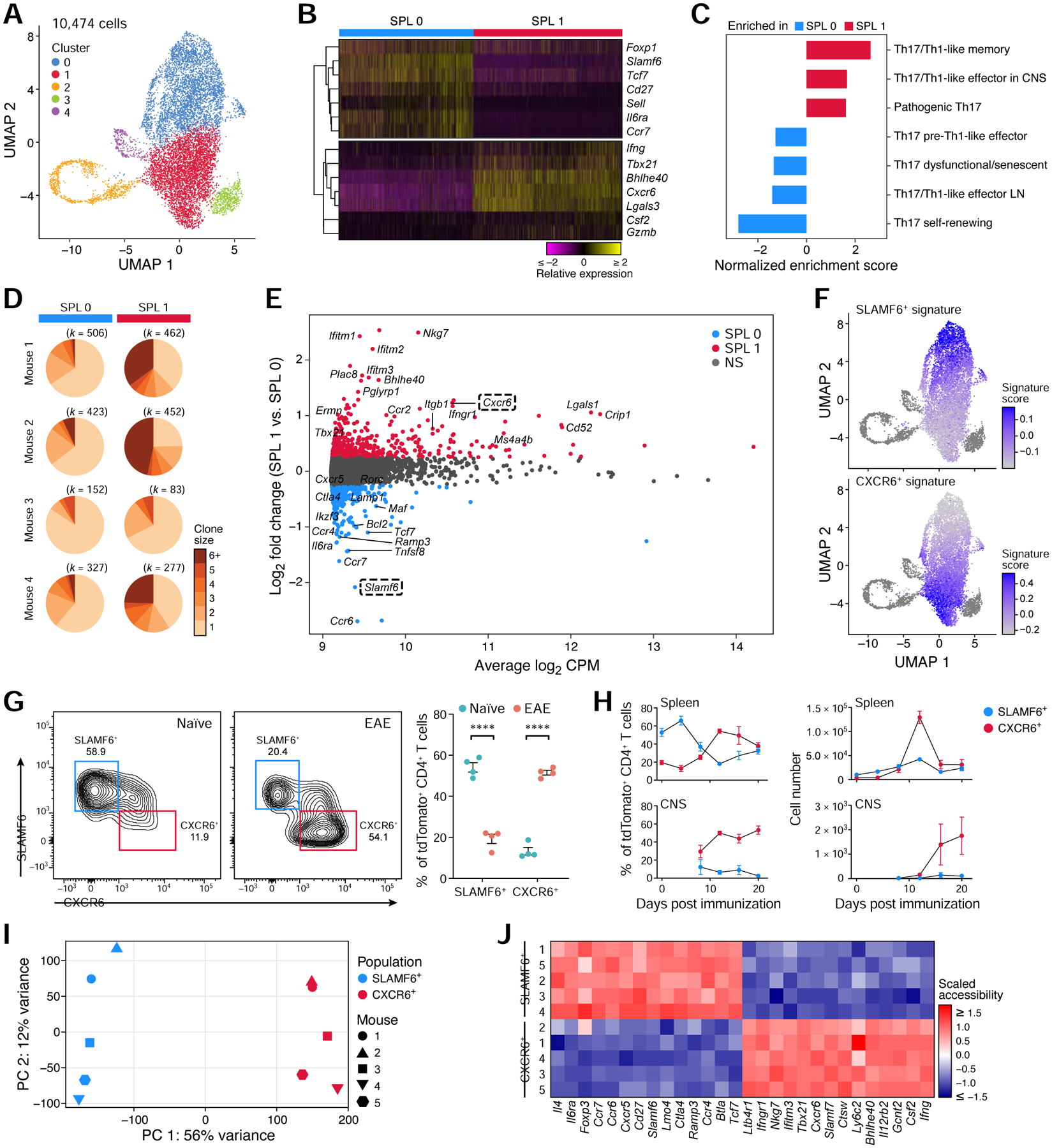
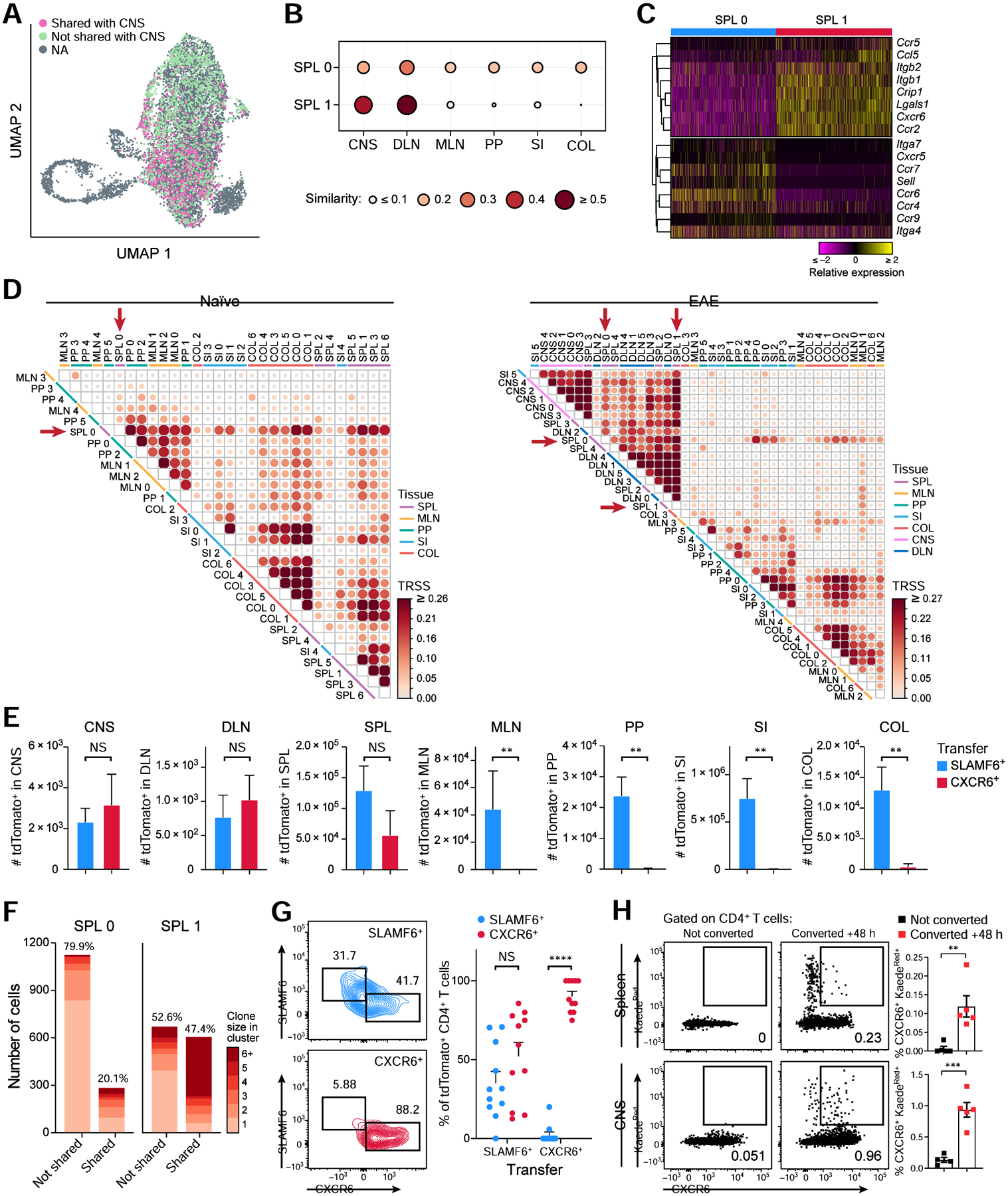
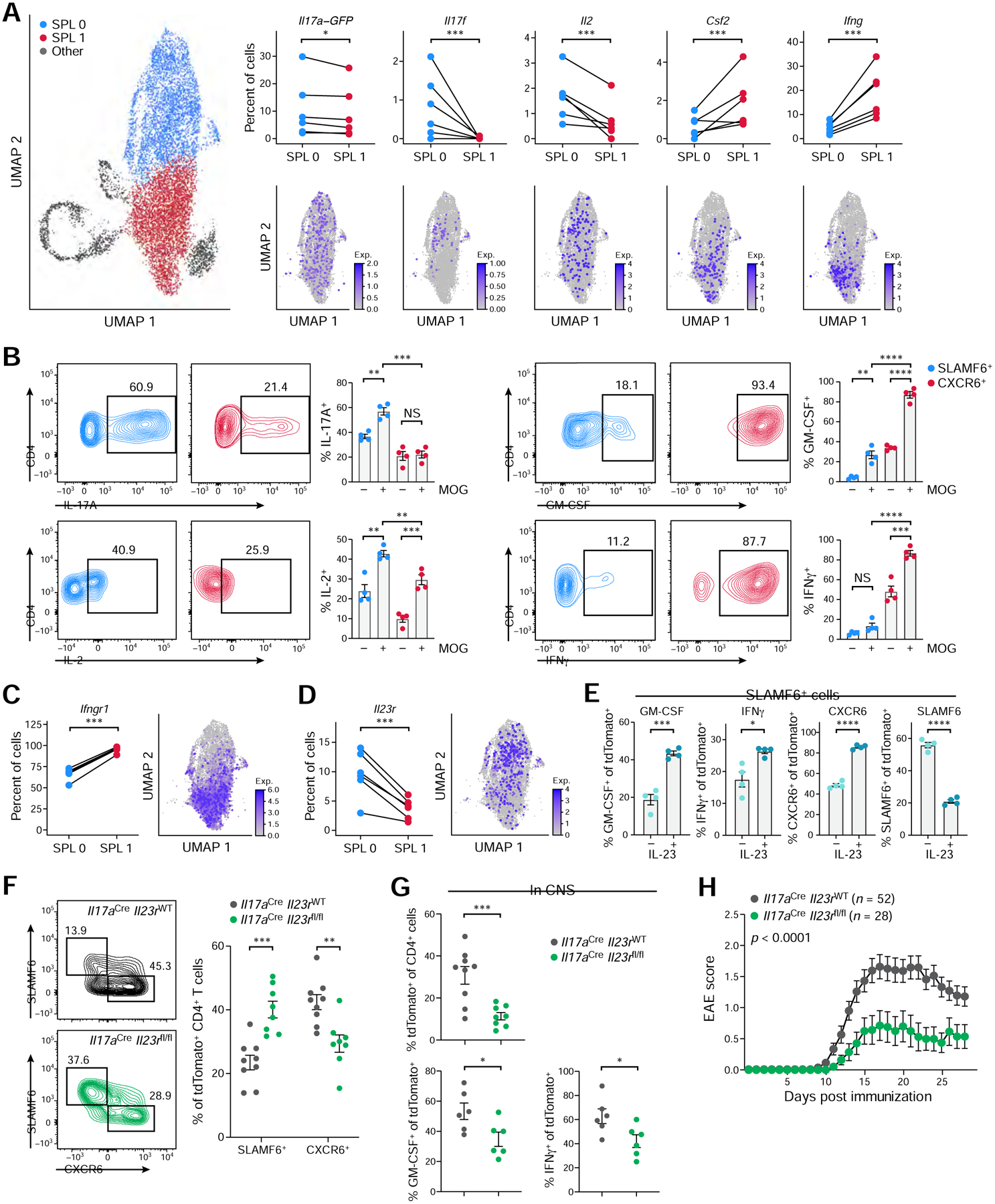
While intestinal Th17 cells are critical for maintaining tissue homeostasis, recent studies have implicated their roles in the development of extra-intestinal autoimmune diseases including multiple sclerosis. However, the mechanisms by which tissue Th17 cells mediate these dichotomous functions remain unknown. Here, we characterized the heterogeneity, plasticity, and migratory phenotypes of tissue Th17 cells in vivo by combined fate mapping with profiling of the transcriptomes and TCR clonotypes of over 84,000 Th17 cells at homeostasis and during CNS autoimmune inflammation. Inter- and intra-organ single-cell analyses revealed a homeostatic, stem-like TCF1+ IL-17+ SLAMF6+ population that traffics to the intestine where it is maintained by the microbiota, providing a ready reservoir for the IL-23-driven generation of encephalitogenic GM-CSF+ IFN-γ+ CXCR6+ T cells. Our study defines a direct in vivo relationship between IL-17+ non-pathogenic and GM-CSF+ and IFN-γ+ pathogenic Th17 populations and provides a mechanism by which homeostatic intestinal Th17 cells direct extra-intestinal autoimmune disease.
Keywords: CNS inflammation; GM-CSF; IFNγ; IL-17; Th17 cells; autoimmunity; fate-mapping; gut-brain axis; multiple sclerosis; stem-like T cells.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.R. was a SAB member of Thermo Fisher Scientific, Neogene Therapeutics, Asimov, and Syros Pharmaceuticals. A.R. is a cofounder of and equity holder in Celsius Therapeutics and an equity holder in Immunitas. V.K.K. is a co-founder and has an ownership interest and is a member of SAB in Celsius Therapeutics, Tizona Therapeutics, and Trishula Therapeutics. V.K.K. is also the chair of the board and has equity interests in Bicara Therpeutics. V.K.K.’s financial interests and conflicts are managed by Brigham and Women’s Hospital and Partners Health Care system. A.R. and O.R.-R. are employees of Genentech (member of the Roche Group) since August and October 2020, respectively. None of these companies provided support for this work. A.R. was an HHMI Investigator while this study was conducted. A. Schnell, M.S., A.R., and V.K.K. are co-inventors on US provisional patent application no. 17/063,617 and no. 62/968,981 filed by the Broad Institute relating to the subject matter of this manuscript. O.R.-R. is a co-inventor on patent applications filed by the Broad Institute relating to single-cell genomics. All other authors declare no competing interests.
Figures
Comment in
-
Th17 cells: from gut homeostasis to CNS pathogenesis.Trends Immunol. 2022 Mar;43(3):167-169. doi: 10.1016/j.it.2022.01.005. Epub 2022 Jan 17. Trends Immunol. 2022. PMID: 35058150 Free PMC article.
References
-
- ARAZI A, RAO DA, BERTHIER CC, DAVIDSON A, LIU Y, HOOVER PJ, CHICOINE A, EISENHAURE TM, JONSSON AH, LI S, LIEB DJ, ZHANG F, SLOWIKOWSKI K, BROWNE EP, NOMA A, SUTHERBY D, STEELMAN S, SMILEK DE, TOSTA P, APRUZZESE W, MASSAROTTI E, DALL’ERA M, PARK M, KAMEN DL, FURIE RA, PAYAN-SCHOBER F, PENDERGRAFT WF 3RD, MCINNIS EA, BUYON JP, PETRI MA, PUTTERMAN C, KALUNIAN KC, WOODLE ES, LEDERER JA, HILDEMAN DA, NUSBAUM C, RAYCHAUDHURI S, KRETZLER M, ANOLIK JH, BRENNER MB, WOFSY D, HACOHEN N, DIAMOND B & ACCELERATING MEDICINES PARTNERSHIP IN, S. L. E. N. 2019. The immune cell landscape in kidneys of patients with lupus nephritis. Nat Immunol, 20, 902–914. - PMC - PubMed
-
- BERER K, GERDES LA, CEKANAVICIUTE E, JIA X, XIAO L, XIA Z, LIU C, KLOTZ L, STAUFFER U, BARANZINI SE, KUMPFEL T, HOHLFELD R, KRISHNAMOORTHY G & WEKERLE H 2017. Gut microbiota from multiple sclerosis patients enables spontaneous autoimmune encephalomyelitis in mice. Proc Natl Acad Sci U S A, 114, 10719–10724. - PMC - PubMed
-
- BERER K, MUES M, KOUTROLOS M, RASBI ZA, BOZIKI M, JOHNER C, WEKERLE H & KRISHNAMOORTHY G 2011. Commensal microbiota and myelin autoantigen cooperate to trigger autoimmune demyelination. Nature, 479, 538–41. - PubMed
-
- BETTELLI E, CARRIER Y, GAO W, KORN T, STROM TB, OUKKA M, WEINER HL & KUCHROO VK 2006. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature, 441, 235–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
