

Microtopographical guidance of macropinocytic signaling patches
- PMID: 34876521
- PMCID: PMC8685668
- DOI: 10.1073/pnas.2110281118
Microtopographical guidance of macropinocytic signaling patches
Abstract
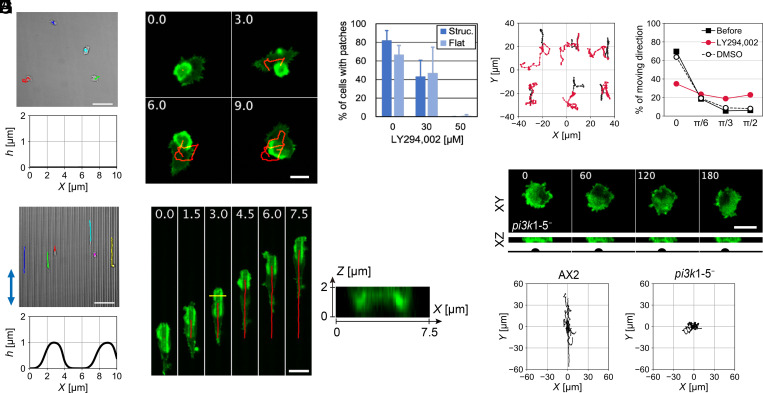
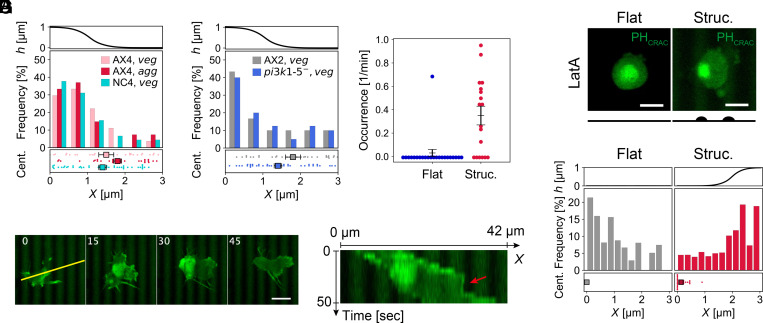
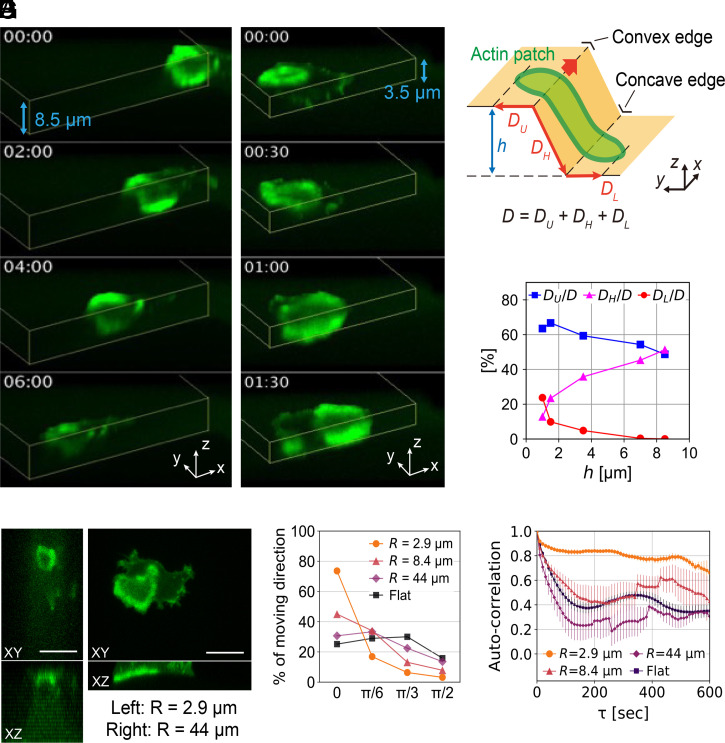
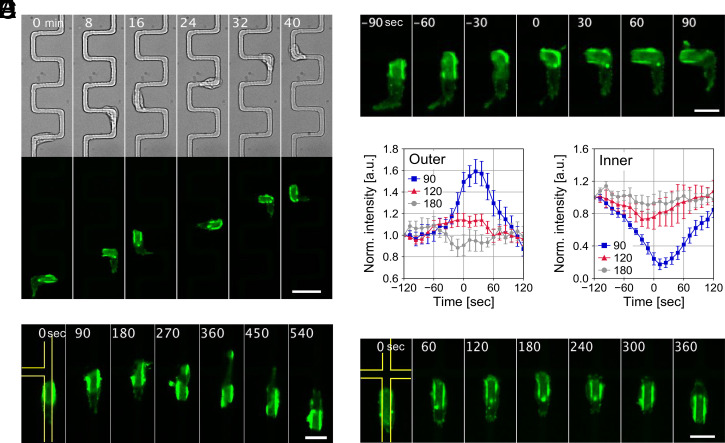
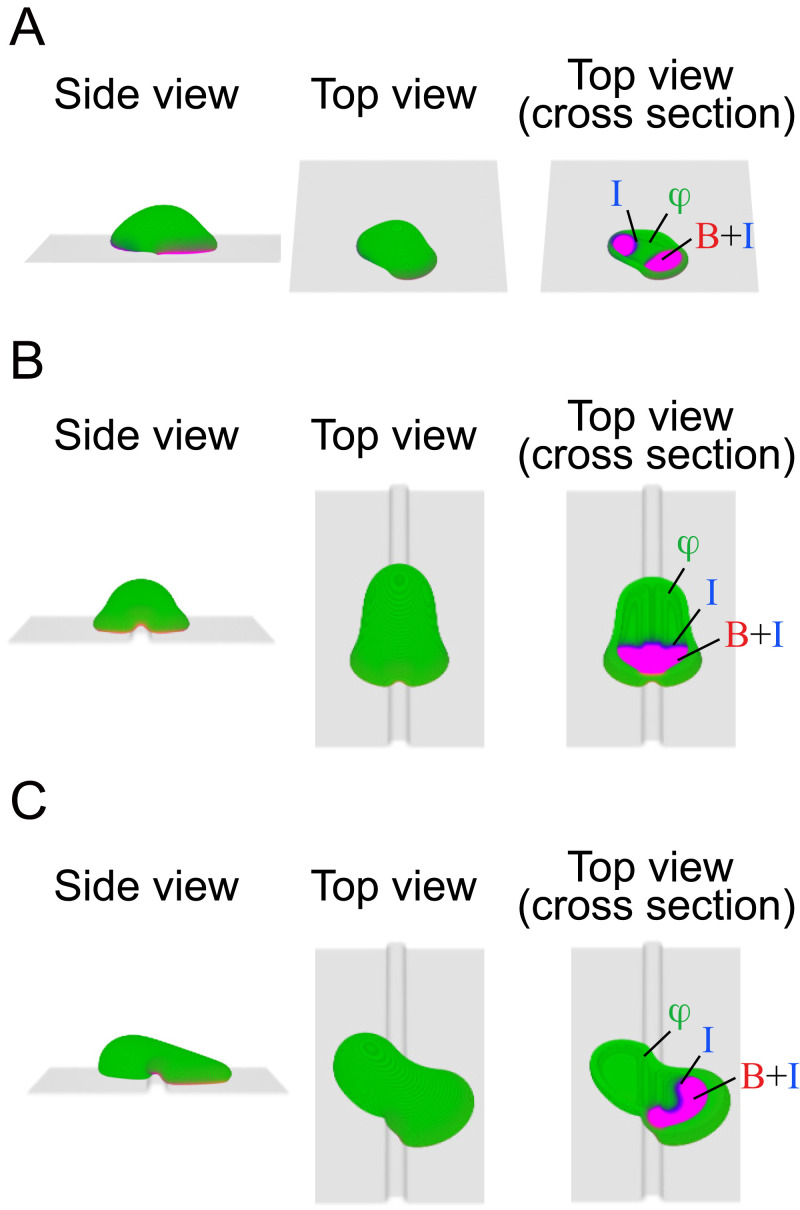
In fast-moving cells such as amoeba and immune cells, dendritic actin filaments are spatiotemporally regulated to shape large-scale plasma membrane protrusions. Despite their importance in migration, as well as in particle and liquid ingestion, how their dynamics are affected by micrometer-scale features of the contact surface is still poorly understood. Here, through quantitative image analysis of Dictyostelium on microfabricated surfaces, we show that there is a distinct mode of topographical guidance directed by the macropinocytic membrane cup. Unlike other topographical guidance known to date that depends on nanometer-scale curvature sensing protein or stress fibers, the macropinocytic membrane cup is driven by the Ras/PI3K/F-actin signaling patch and its dependency on the micrometer-scale topographical features, namely PI3K/F-actin-independent accumulation of Ras-GTP at the convex curved surface, PI3K-dependent patch propagation along the convex edge, and its actomyosin-dependent constriction at the concave edge. Mathematical model simulations demonstrate that the topographically dependent initiation, in combination with the mutually defining patch patterning and the membrane deformation, gives rise to the topographical guidance. Our results suggest that the macropinocytic cup is a self-enclosing structure that can support liquid ingestion by default; however, in the presence of structured surfaces, it is directed to faithfully trace bent and bifurcating ridges for particle ingestion and cell guidance.
Keywords: actin waves; cell migration; contact guidance; macropinocytosis; topography.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Reig G., Pulgar E., Concha M. L., Cell migration: From tissue culture to embryos. Development 141, 1999–2013 (2014). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
