

Diurnal metabolic control in cyanobacteria requires perception of second messenger signaling molecule c-di-AMP by the carbon control protein SbtB
- PMID: 34878830
- PMCID: PMC8654305
- DOI: 10.1126/sciadv.abk0568
Diurnal metabolic control in cyanobacteria requires perception of second messenger signaling molecule c-di-AMP by the carbon control protein SbtB
Abstract
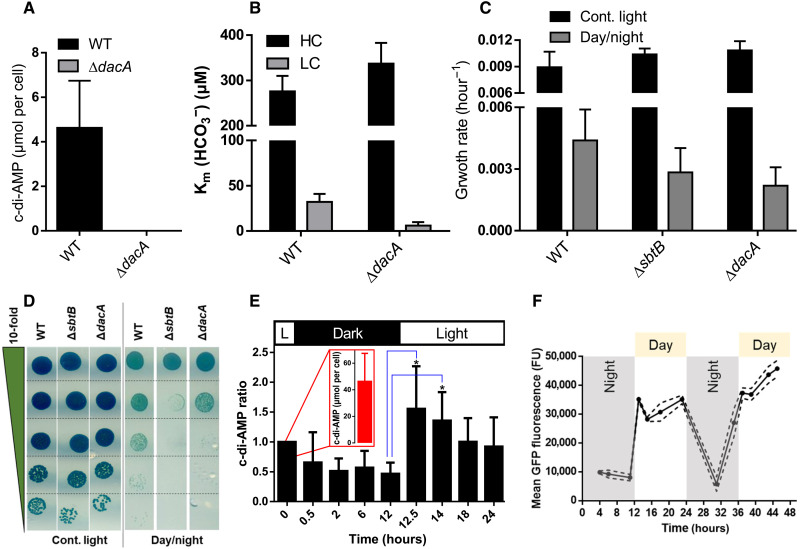
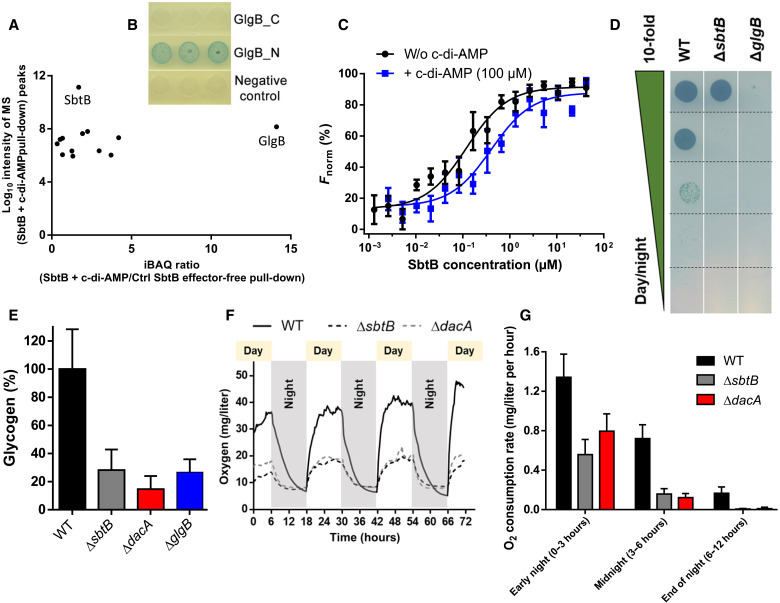
Because of their photosynthesis-dependent lifestyle, cyanobacteria evolved sophisticated regulatory mechanisms to adapt to oscillating day-night metabolic changes. How they coordinate the metabolic switch between autotrophic and glycogen-catabolic metabolism in light and darkness is poorly understood. Recently, c-di-AMP has been implicated in diurnal regulation, but its mode of action remains elusive. To unravel the signaling functions of c-di-AMP in cyanobacteria, we isolated c-di-AMP receptor proteins. Thereby, the carbon-sensor protein SbtB was identified as a major c-di-AMP receptor, which we confirmed biochemically and by x-ray crystallography. In search for the c-di-AMP signaling function of SbtB, we found that both SbtB and c-di-AMP cyclase–deficient mutants showed reduced diurnal growth and that c-di-AMP–bound SbtB interacts specifically with the glycogen-branching enzyme GlgB. Accordingly, both mutants displayed impaired glycogen synthesis during the day and impaired nighttime survival. Thus, the pivotal role of c-di-AMP in day-night acclimation can be attributed to SbtB-mediated regulation of glycogen metabolism.
Figures


Similar articles
-
Carbon signaling protein SbtB possesses atypical redox-regulated apyrase activity to facilitate regulation of bicarbonate transporter SbtA.Proc Natl Acad Sci U S A. 2023 Feb 21;120(8):e2205882120. doi: 10.1073/pnas.2205882120. Epub 2023 Feb 17. Proc Natl Acad Sci U S A. 2023. PMID: 36800386 Free PMC article.
-
Roles of second messengers in the regulation of cyanobacterial physiology: the carbon-concentrating mechanism and beyond.Microlife. 2023 Feb 23;4:uqad008. doi: 10.1093/femsml/uqad008. eCollection 2023. Microlife. 2023. PMID: 37223741 Free PMC article. Review.
-
PII-like signaling protein SbtB links cAMP sensing with cyanobacterial inorganic carbon response.Proc Natl Acad Sci U S A. 2018 May 22;115(21):E4861-E4869. doi: 10.1073/pnas.1803790115. Epub 2018 May 7. Proc Natl Acad Sci U S A. 2018. PMID: 29735650 Free PMC article.
-
The impact of the cyanobacterial carbon-regulator protein SbtB and of the second messengers cAMP and c-di-AMP on CO2 -dependent gene expression.New Phytol. 2022 Jun;234(5):1801-1816. doi: 10.1111/nph.18094. Epub 2022 Apr 9. New Phytol. 2022. PMID: 35285042
-
Replenishing the cyclic-di-AMP pool: regulation of diadenylate cyclase activity in bacteria.Curr Genet. 2016 Nov;62(4):731-738. doi: 10.1007/s00294-016-0600-8. Epub 2016 Apr 13. Curr Genet. 2016. PMID: 27074767 Review.
Cited by
-
Sustained Control of Pyruvate Carboxylase by the Essential Second Messenger Cyclic di-AMP in Bacillus subtilis.mBio. 2021 Feb 22;13(1):e0360221. doi: 10.1128/mbio.03602-21. Epub 2022 Feb 8. mBio. 2021. PMID: 35130724 Free PMC article.
-
Carbon signaling protein SbtB possesses atypical redox-regulated apyrase activity to facilitate regulation of bicarbonate transporter SbtA.Proc Natl Acad Sci U S A. 2023 Feb 21;120(8):e2205882120. doi: 10.1073/pnas.2205882120. Epub 2023 Feb 17. Proc Natl Acad Sci U S A. 2023. PMID: 36800386 Free PMC article.
-
ComFB, a new widespread family of c-di-NMP receptor proteins.bioRxiv [Preprint]. 2024 Nov 10:2024.11.10.622515. doi: 10.1101/2024.11.10.622515. bioRxiv. 2024. PMID: 39574629 Free PMC article. Preprint.
-
Bacterial cell volume regulation and the importance of cyclic di-AMP.Microbiol Mol Biol Rev. 2024 Jun 27;88(2):e0018123. doi: 10.1128/mmbr.00181-23. Epub 2024 Jun 10. Microbiol Mol Biol Rev. 2024. PMID: 38856222 Free PMC article. Review.
-
cAMP Signaling in Cancer: A PKA-CREB and EPAC-Centric Approach.Cells. 2022 Jun 24;11(13):2020. doi: 10.3390/cells11132020. Cells. 2022. PMID: 35805104 Free PMC article. Review.
References
-
- Wan N., DeLorenzo D. M., He L., You L., Immethun C. M., Wang G., Baidoo E. E. K., Hollinshead W., Keasling J. D., Moon T. S., Tang Y. J., Cyanobacterial carbon metabolism: Fluxome plasticity and oxygen dependence. Biotechnol. Bioeng. 114, 1593–1602 (2017). - PubMed
-
- Makowka A., Nichelmann L., Schulze D., Spengler K., Wittmann C., Forchhammer K., Gutekunst K., Glycolytic Shunts Replenish the Calvin–Benson–Bassham Cycle as Anaplerotic Reactions in Cyanobacteria. Mol. Plant 13, 471–482 (2020). - PubMed
LinkOut - more resources
Full Text Sources
