

A secretory phospholipase D hydrolyzes phosphatidylcholine to suppress rice heading time
- PMID: 34879072
- PMCID: PMC8654219
- DOI: 10.1371/journal.pgen.1009905
A secretory phospholipase D hydrolyzes phosphatidylcholine to suppress rice heading time
Abstract
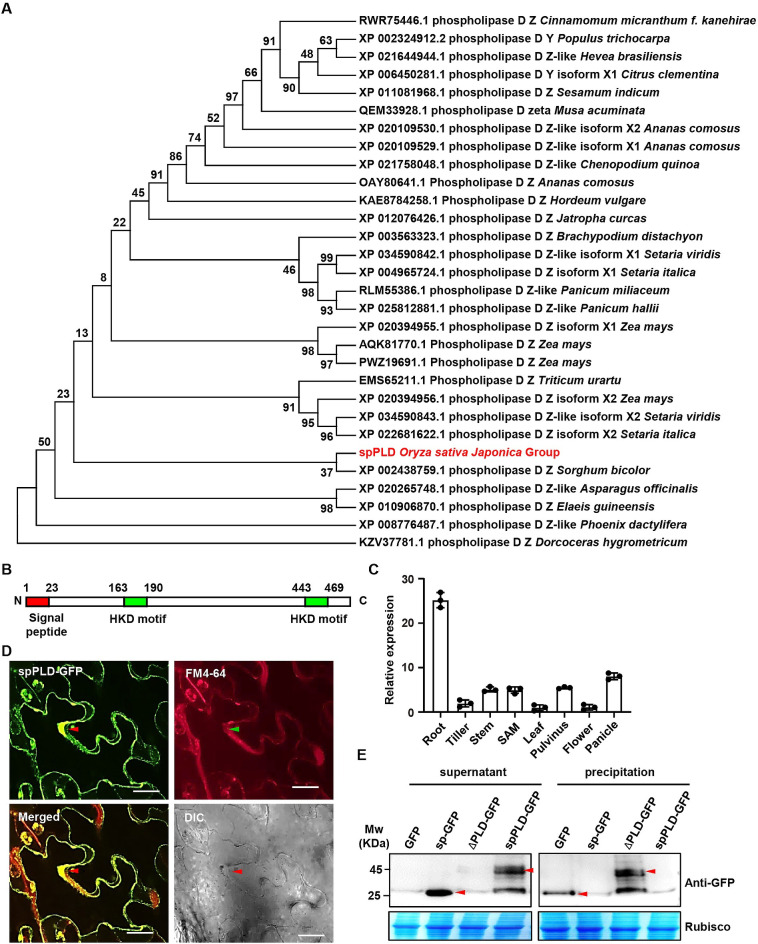
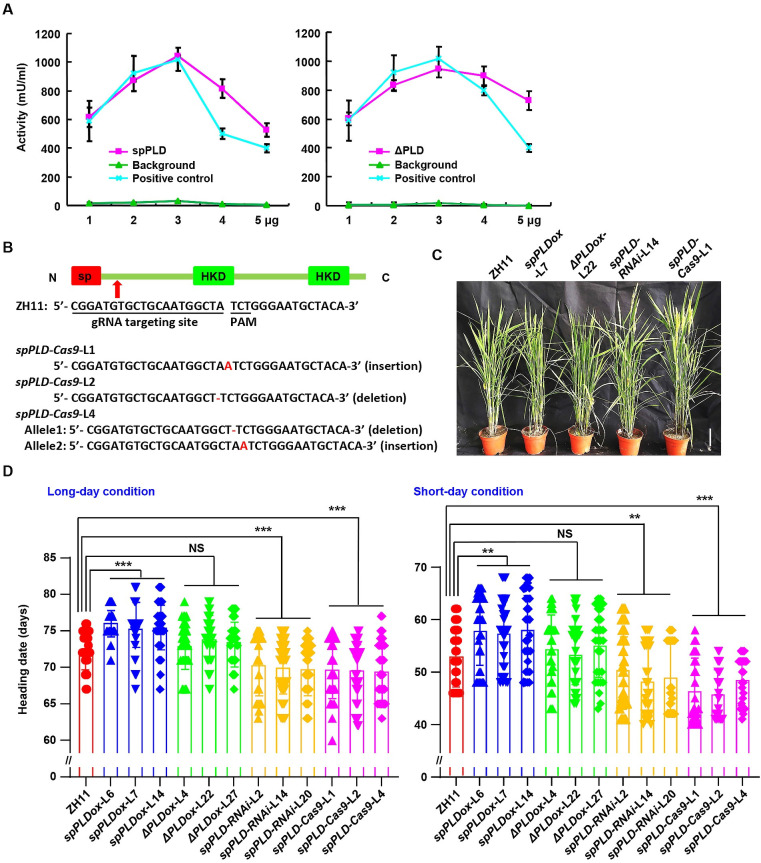
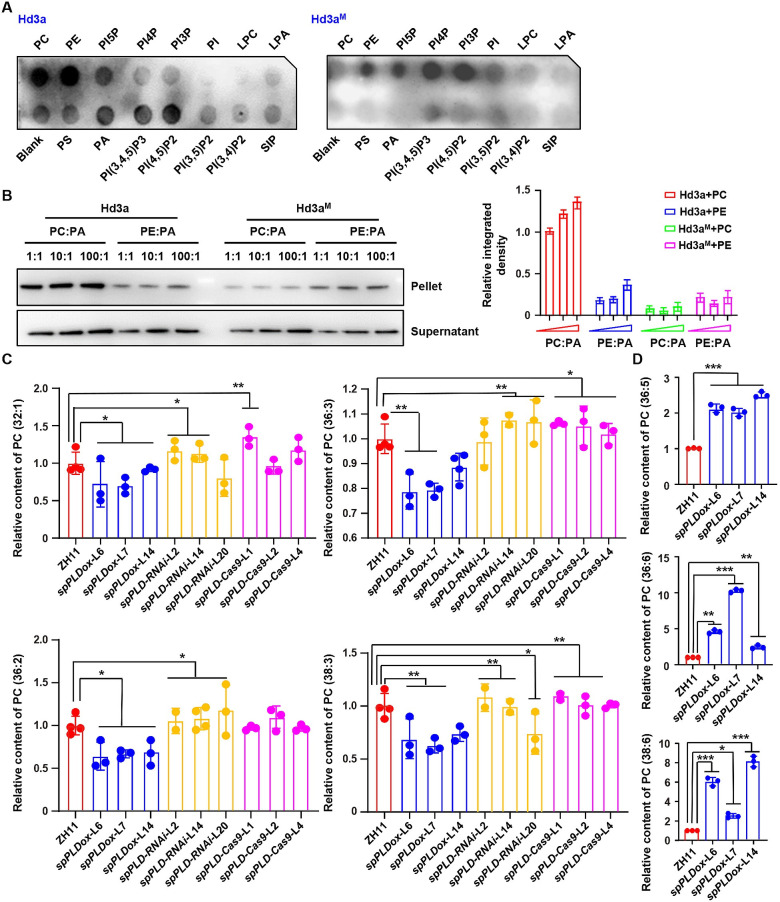
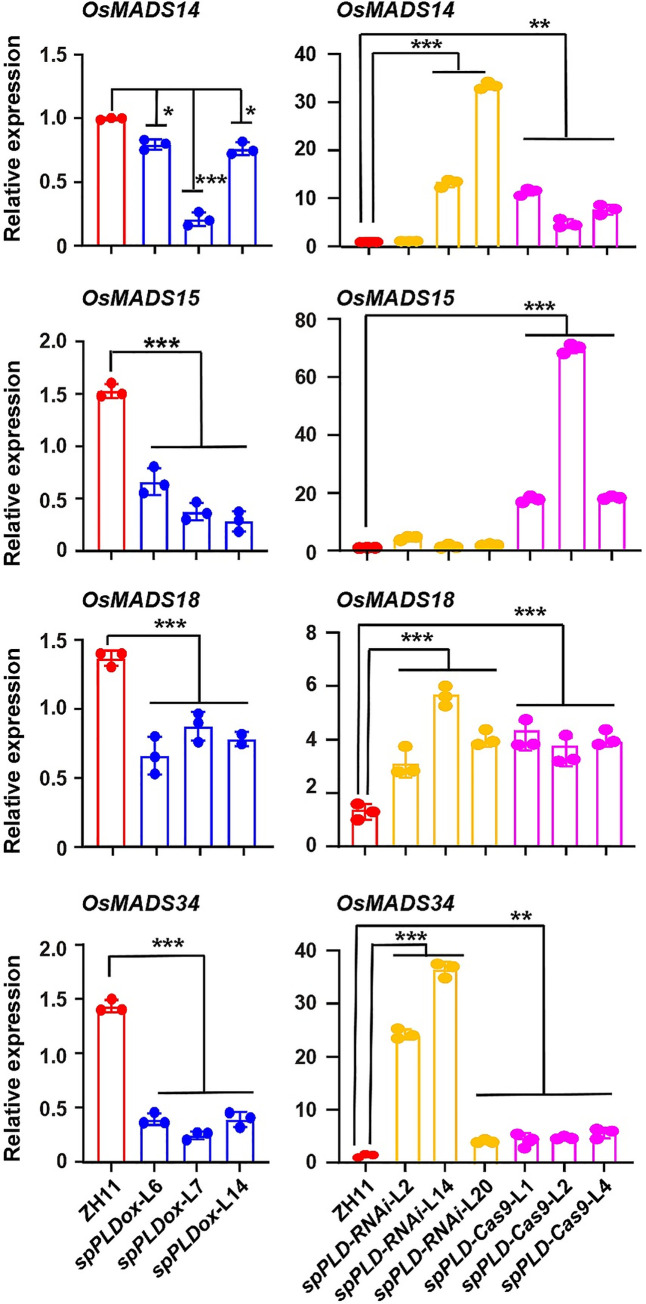
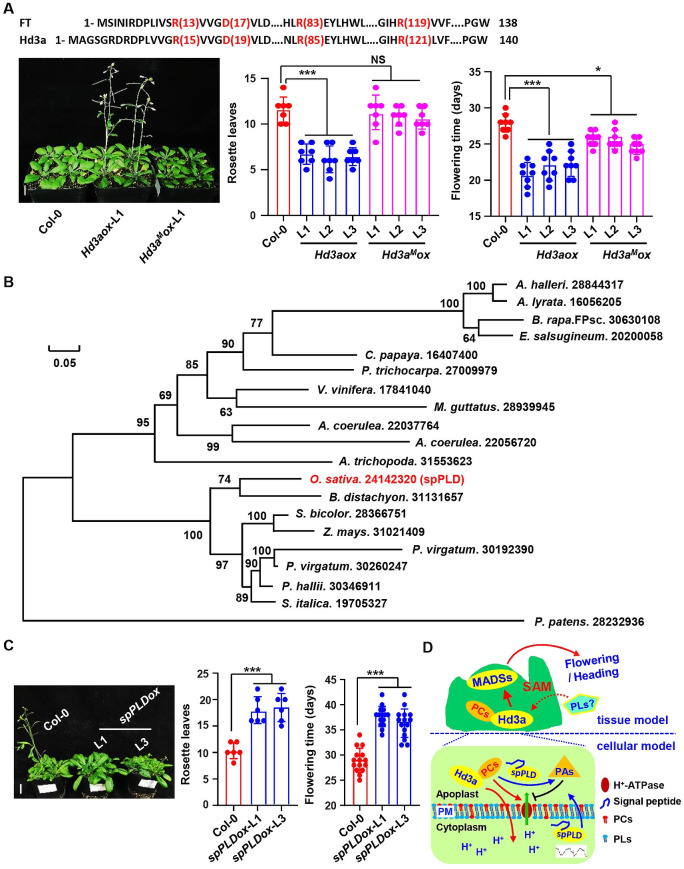
Phospholipase D (PLD) hydrolyzes membrane phospholipids and is crucial in various physiological processes and transduction of different signals. Secretory phospholipases play important roles in mammals, however, whose functions in plants remain largely unknown. We previously identified a rice secretory PLD (spPLD) that harbors a signal peptide and here we reported the secretion and function of spPLD in rice heading time regulation. Subcellular localization analysis confirmed the signal peptide is indispensable for spPLD secretion into the extracellular spaces, where spPLD hydrolyzes substrates. spPLD overexpression results in delayed heading time which is dependent on its secretory character, while suppression or deficiency of spPLD led to the early heading of rice under both short-day and long-day conditions, which is consistent with that spPLD overexpression/suppression indeed led to the reduced/increased Hd3a/RFT1 (Arabidopsis Flowing Locus T homolog) activities. Interestingly, rice Hd3a and RFT1 bind to phosphatidylcholines (PCs) and a further analysis by lipidomic approach using mass spectrometry revealed the altered phospholipids profiles in shoot apical meristem, particularly the PC species, under altered spPLD expressions. These results indicate the significance of secretory spPLD and help to elucidate the regulatory network of rice heading time.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
