

NEAT1 is essential for metabolic changes that promote breast cancer growth and metastasis
- PMID: 34879239
- PMCID: PMC8813003
- DOI: 10.1016/j.cmet.2021.11.011
NEAT1 is essential for metabolic changes that promote breast cancer growth and metastasis
Abstract
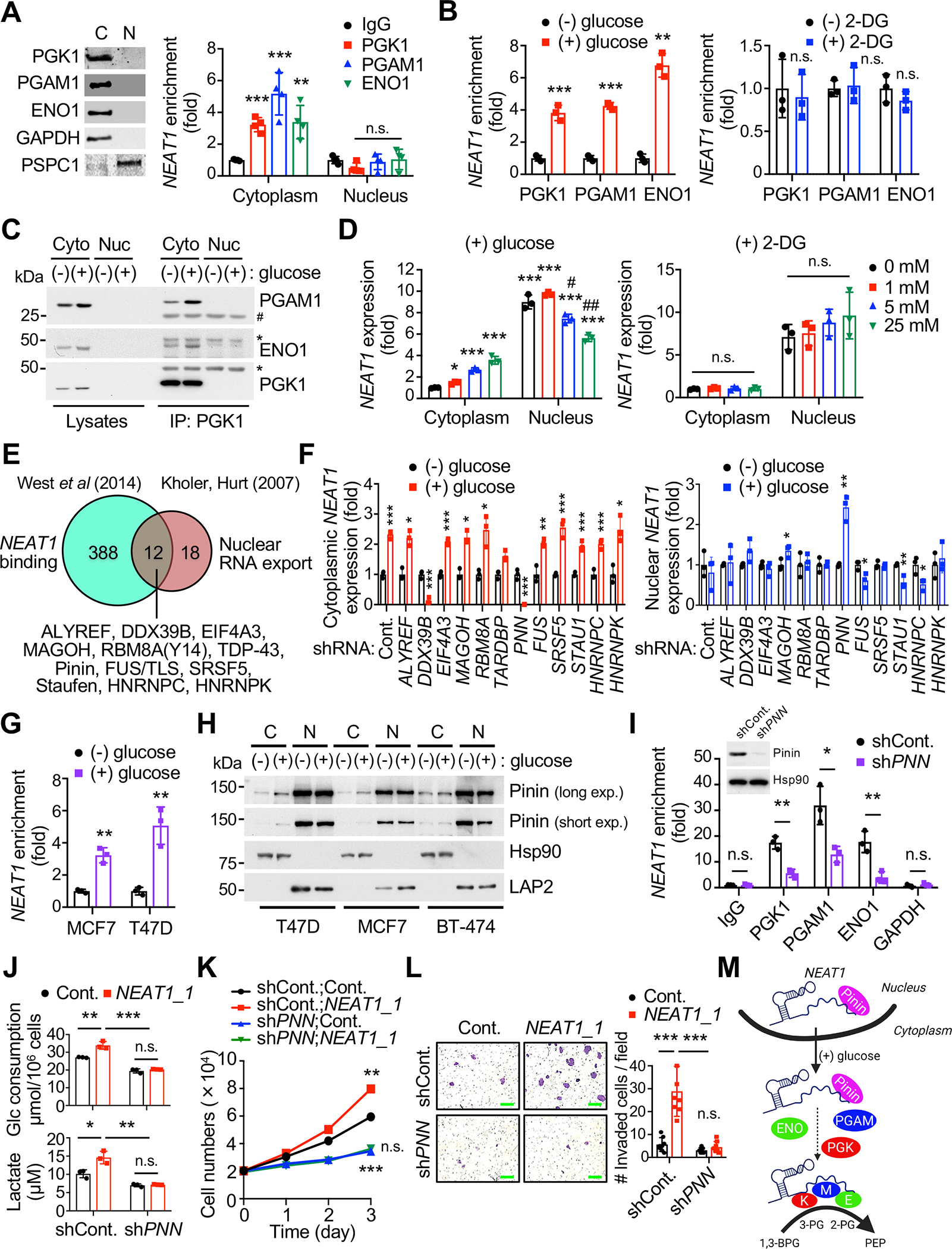
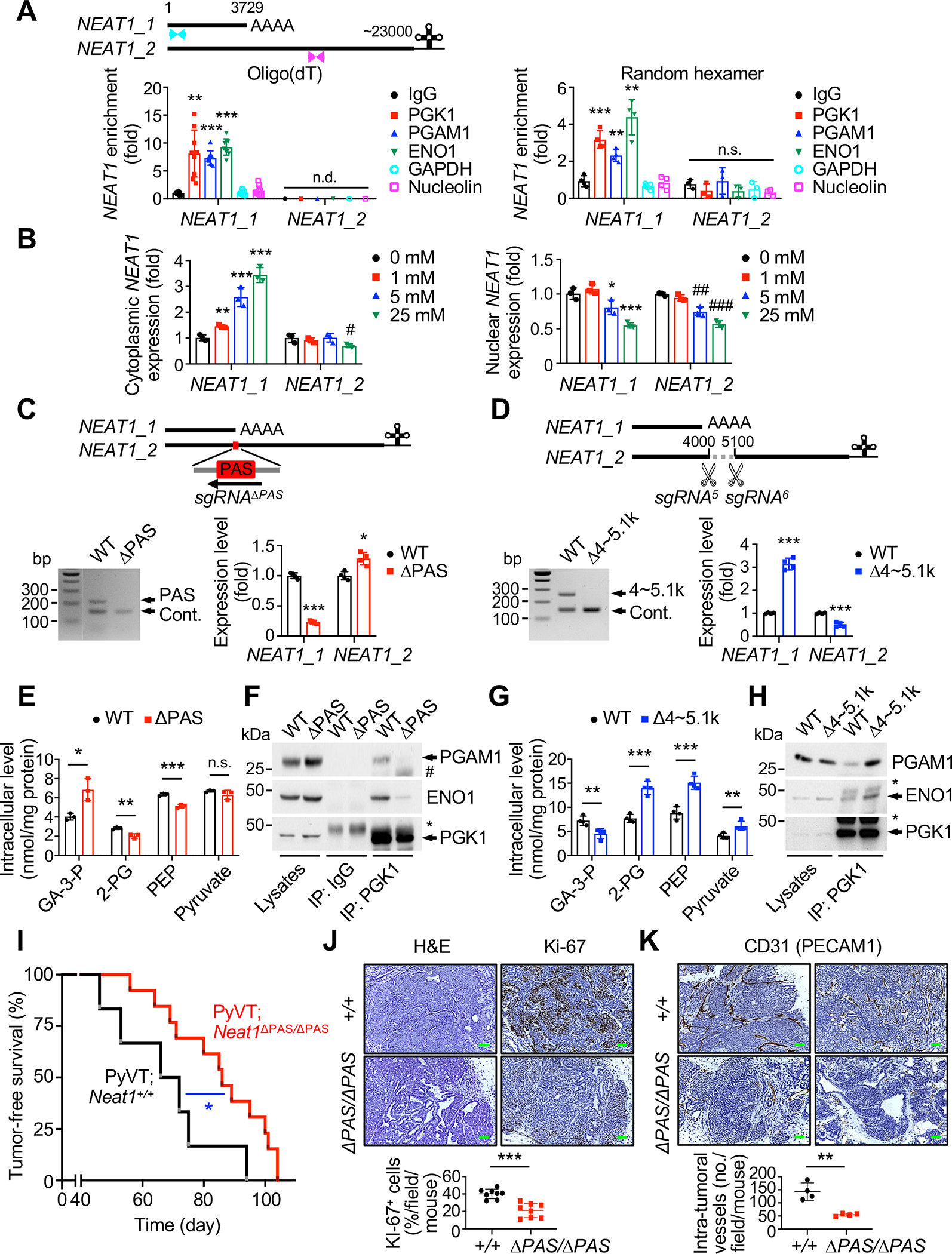
Accelerated glycolysis is the main metabolic change observed in cancer, but the underlying molecular mechanisms and their role in cancer progression remain poorly understood. Here, we show that the deletion of the long noncoding RNA (lncRNA) Neat1 in MMTV-PyVT mice profoundly impairs tumor initiation, growth, and metastasis, specifically switching off the penultimate step of glycolysis. Mechanistically, NEAT1 directly binds and forms a scaffold bridge for the assembly of PGK1/PGAM1/ENO1 complexes and thereby promotes substrate channeling for high and efficient glycolysis. Notably, NEAT1 is upregulated in cancer patients and correlates with high levels of these complexes, and genetic and pharmacological blockade of penultimate glycolysis ablates NEAT1-dependent tumorigenesis. Finally, we demonstrate that Pinin mediates glucose-stimulated nuclear export of NEAT1, through which it exerts isoform-specific and paraspeckle-independent functions. These findings establish a direct role for NEAT1 in regulating tumor metabolism, provide new insights into the Warburg effect, and identify potential targets for therapy.
Keywords: ENO1; NEAT1; PGAM1; PGK1; Pinin; Warburg effect; aerobic glycolysis; breast cancer; long noncoding RNA; tumor metabolism.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures





References
-
- Adriaens C, Standaert L, Barra J, Latil M, Verfaillie A, Kalev P, Boeckx B, Wijnhoven PW, Radaelli E, Vermi W, et al. (2016). p53 induces formation of NEAT1 lncRNA-containing paraspeckles that modulate replication stress response and chemosensitivity. Nat Med 22, 861–868. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
