

Stage-specific regulation of DNA methylation by TET enzymes during human cardiac differentiation
- PMID: 34879277
- PMCID: PMC11229417
- DOI: 10.1016/j.celrep.2021.110095
Stage-specific regulation of DNA methylation by TET enzymes during human cardiac differentiation
Abstract
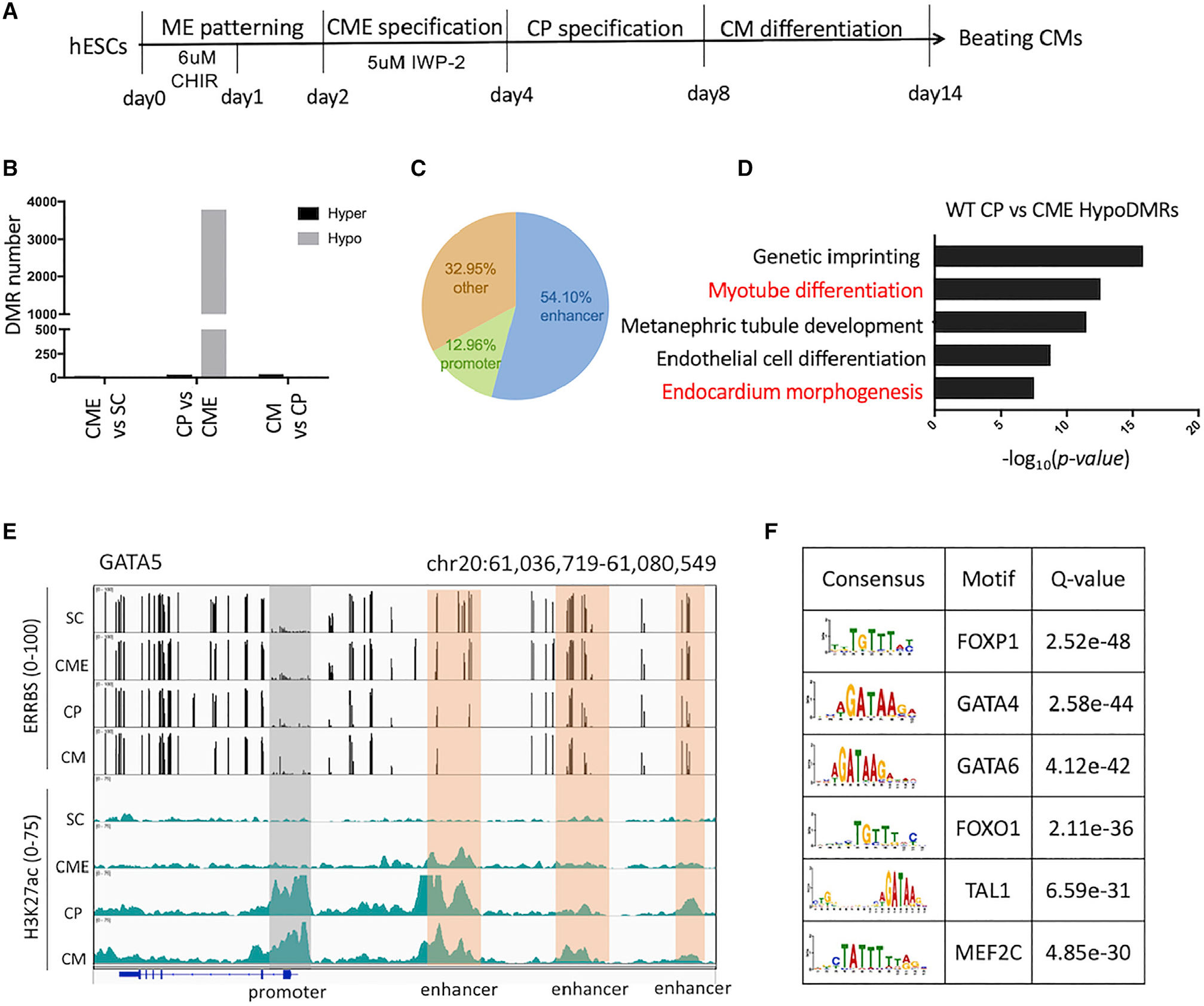
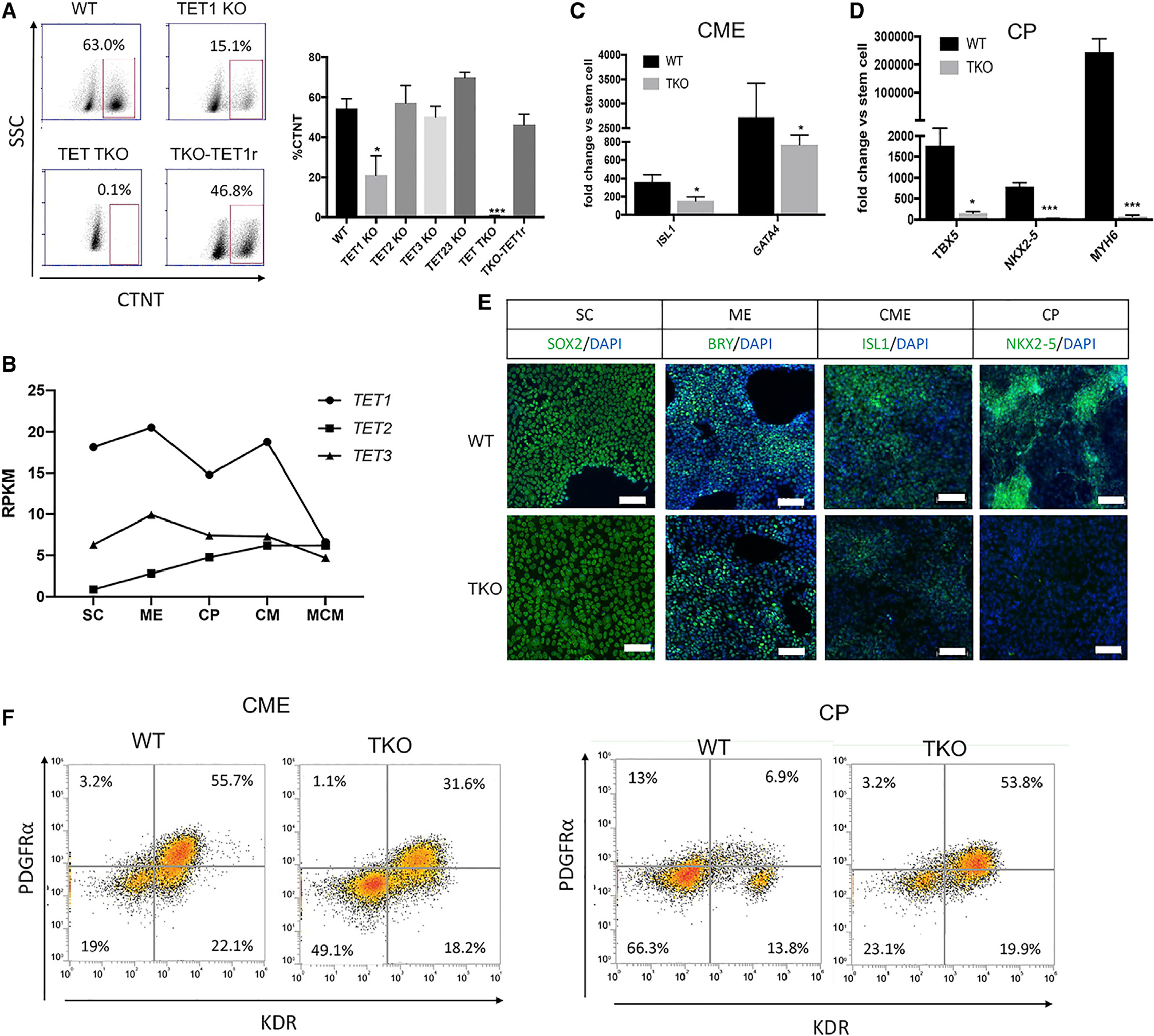
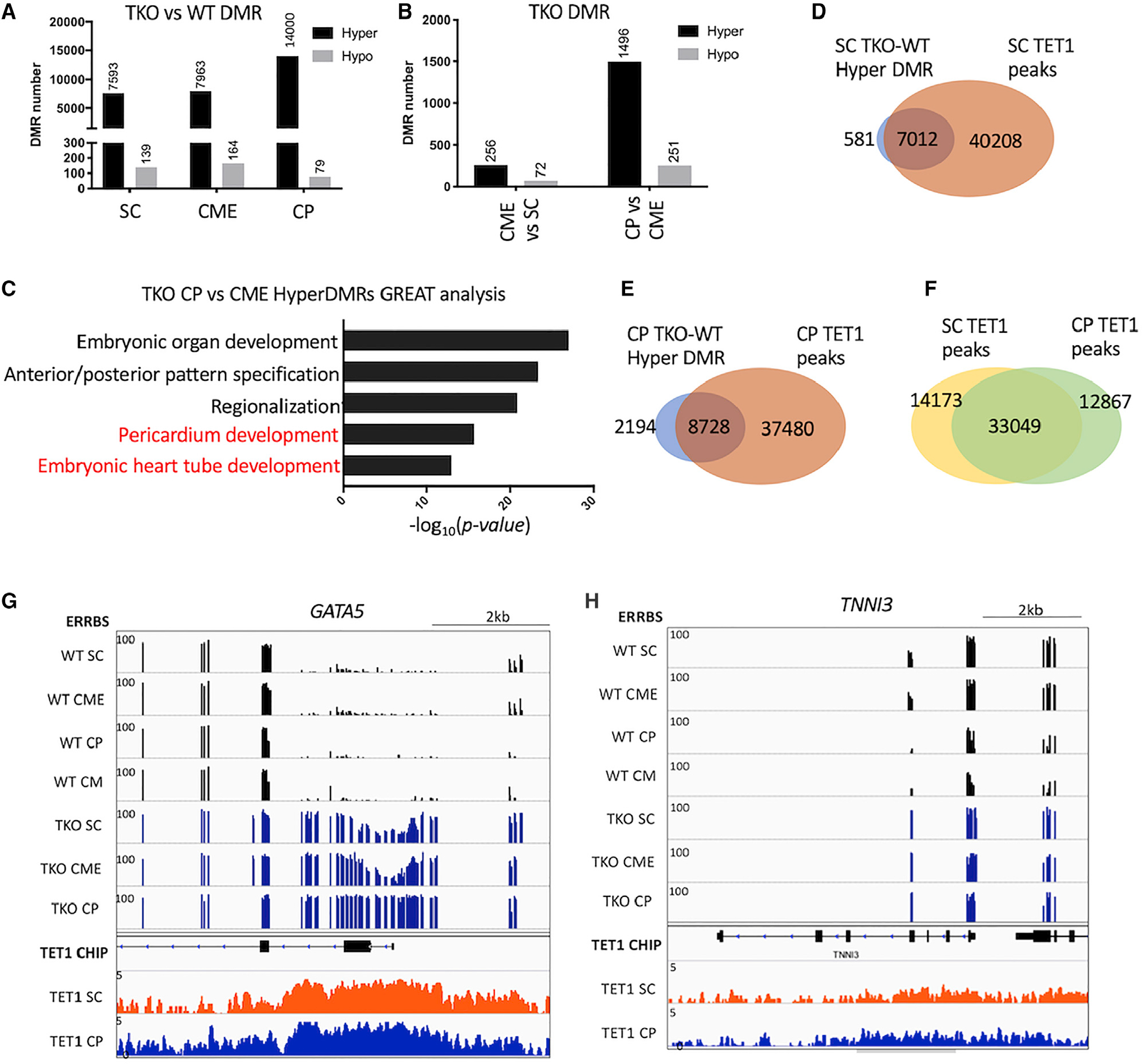
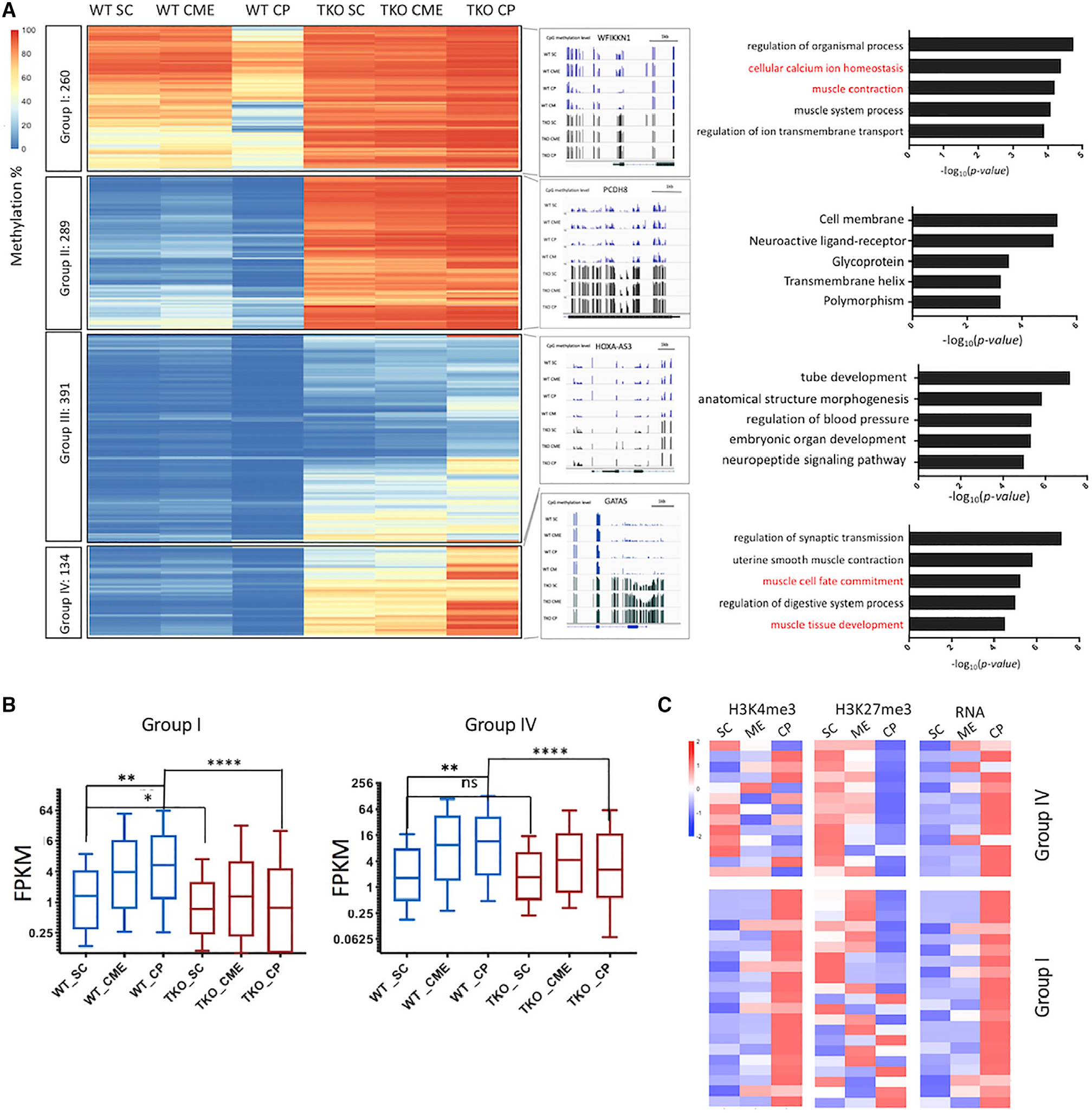
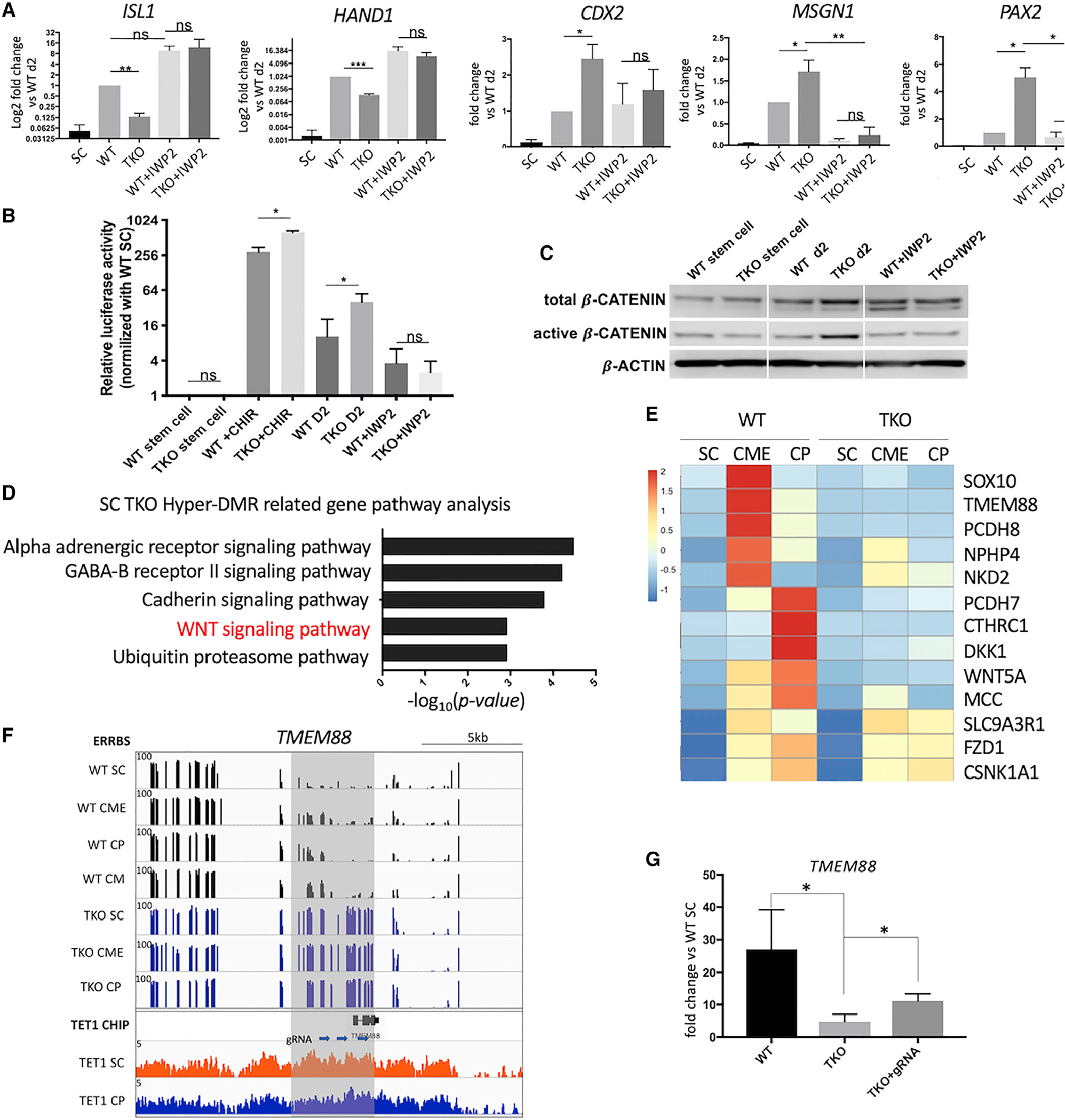
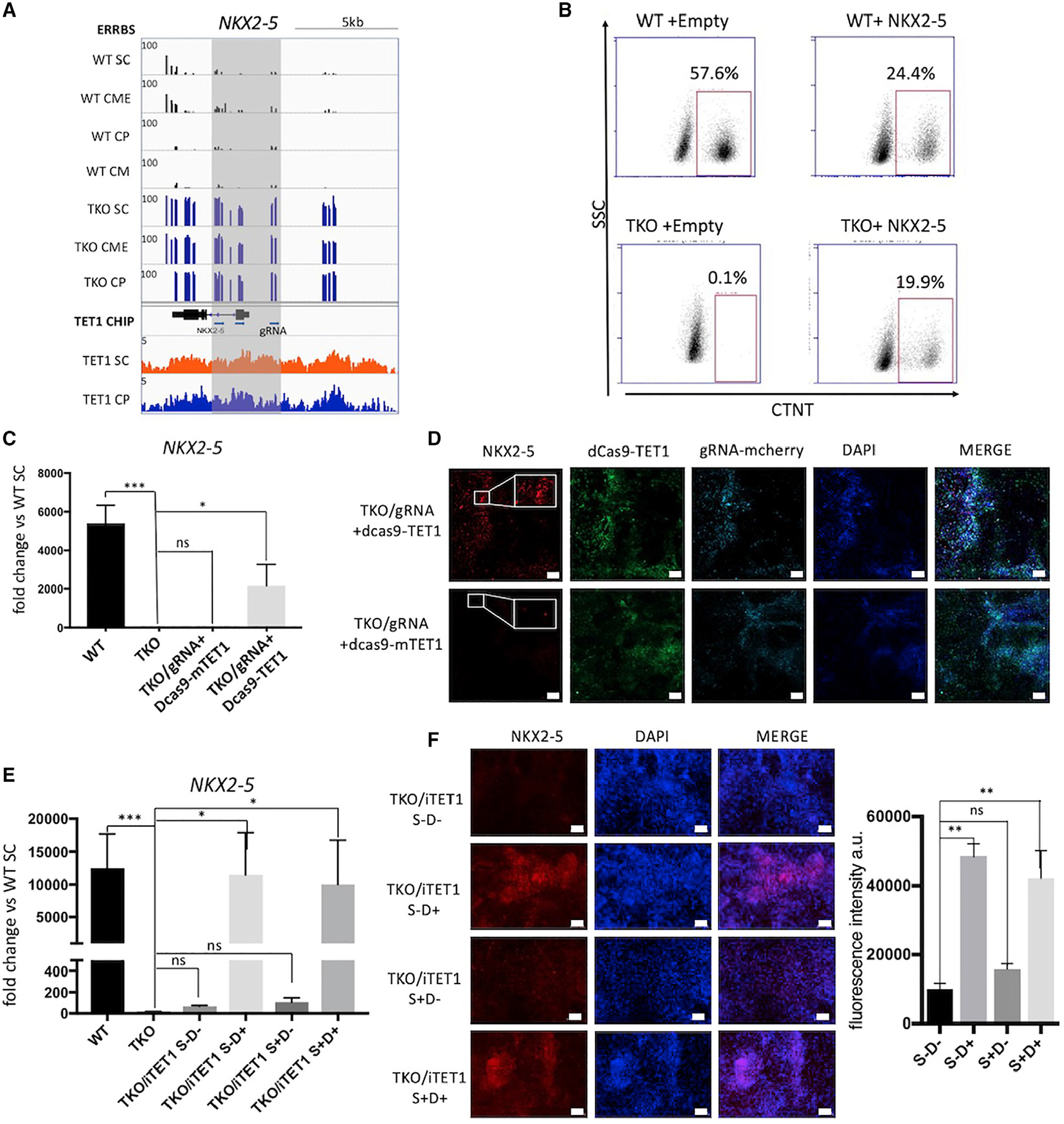
Changes in DNA methylation are associated with normal cardiogenesis, whereas altered methylation patterns can occur in congenital heart disease. Ten-eleven translocation (TET) enzymes oxidize 5-methylcytosine (5mC) and promote locus-specific DNA demethylation. Here, we characterize stage-specific methylation dynamics and the function of TETs during human cardiomyocyte differentiation. Human embryonic stem cells (hESCs) in which all three TET genes are inactivated fail to generate cardiomyocytes (CMs), with altered mesoderm patterning and defective cardiac progenitor specification. Genome-wide methylation analysis shows TET knockout causes promoter hypermethylation of genes encoding WNT inhibitors, leading to hyperactivated WNT signaling and defects in cardiac mesoderm patterning. TET activity is also needed to maintain hypomethylated status and expression of NKX2-5 for subsequent cardiac progenitor specification. Finally, loss of TETs causes a set of cardiac structural genes to fail to be demethylated at the cardiac progenitor stage. Our data demonstrate key roles for TET proteins in controlling methylation dynamics at sequential steps during human cardiac development.
Keywords: NKX2-5; TMEM88; WNT; cardiogenesis; epigenomics; hESCs.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Similar articles
-
TET-mediated DNA demethylation plays an important role in arsenic-induced HBE cells oxidative stress via regulating promoter methylation of OGG1 and GSTP1.Toxicol In Vitro. 2021 Apr;72:105075. doi: 10.1016/j.tiv.2020.105075. Epub 2021 Jan 1. Toxicol In Vitro. 2021. PMID: 33388378
-
Roles and Regulations of TET Enzymes in Solid Tumors.Trends Cancer. 2021 Jul;7(7):635-646. doi: 10.1016/j.trecan.2020.12.011. Epub 2021 Jan 16. Trends Cancer. 2021. PMID: 33468438 Review.
-
TET-mediated DNA demethylation controls gastrulation by regulating Lefty-Nodal signalling.Nature. 2016 Oct 27;538(7626):528-532. doi: 10.1038/nature20095. Epub 2016 Oct 19. Nature. 2016. PMID: 27760115
-
Mechanisms that regulate the activities of TET proteins.Cell Mol Life Sci. 2022 Jun 15;79(7):363. doi: 10.1007/s00018-022-04396-x. Cell Mol Life Sci. 2022. PMID: 35705880 Free PMC article. Review.
-
TET proteins safeguard bivalent promoters from de novo methylation in human embryonic stem cells.Nat Genet. 2018 Jan;50(1):83-95. doi: 10.1038/s41588-017-0002-y. Epub 2017 Dec 4. Nat Genet. 2018. PMID: 29203910 Free PMC article.
Cited by
-
The Role of Selected Epigenetic Pathways in Cardiovascular Diseases as a Potential Therapeutic Target.Int J Mol Sci. 2023 Sep 6;24(18):13723. doi: 10.3390/ijms241813723. Int J Mol Sci. 2023. PMID: 37762023 Free PMC article. Review.
-
GATA6 regulates WNT and BMP programs to pattern precardiac mesoderm during the earliest stages of human cardiogenesis.bioRxiv [Preprint]. 2025 Jan 11:2024.07.09.602666. doi: 10.1101/2024.07.09.602666. bioRxiv. 2025. Update in: Elife. 2025 Mar 13;13:RP100797. doi: 10.7554/eLife.100797. PMID: 39026742 Free PMC article. Updated. Preprint.
-
Epigenetics in Congenital Heart Disease.J Am Heart Assoc. 2022 Apr 5;11(7):e025163. doi: 10.1161/JAHA.121.025163. Epub 2022 Mar 29. J Am Heart Assoc. 2022. PMID: 35348004 Free PMC article. Review.
-
Identification of miR-20b-5p as an inhibitory regulator in cardiac differentiation via TET2 and DNA hydroxymethylation.Clin Epigenetics. 2024 Mar 15;16(1):42. doi: 10.1186/s13148-024-01653-7. Clin Epigenetics. 2024. PMID: 38491513 Free PMC article.
-
Nuclear Control of Vascular Smooth Muscle Cell Plasticity during Vascular Remodeling.Am J Pathol. 2024 Apr;194(4):525-538. doi: 10.1016/j.ajpath.2023.09.013. Epub 2023 Oct 10. Am J Pathol. 2024. PMID: 37820925 Free PMC article. Review.
References
-
- Almeida RD, Loose M, Sottile V, Matsa E, Denning C, Young L, Johnson AD, Gering M, and Ruzov A (2012). 5-hydroxymethyl-cytosine enrichment of non-committed cells is not a universal feature of vertebrate development. Epigenetics 7, 383–389. - PubMed
-
- Dai HQ, Wang BA, Yang L, Chen JJ, Zhu GC, Sun ML, Ge H, Wang R, Chapman DL, Tang F, et al. (2016). TET-mediated DNA demethylation controls gastrulation by regulating Lefty-Nodal signalling. Nature 538, 528–532. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
