

Nucleolar stress controls mutant Huntington toxicity and monitors Huntington's disease progression
- PMID: 34880223
- PMCID: PMC8655027
- DOI: 10.1038/s41419-021-04432-x
Nucleolar stress controls mutant Huntington toxicity and monitors Huntington's disease progression
Abstract
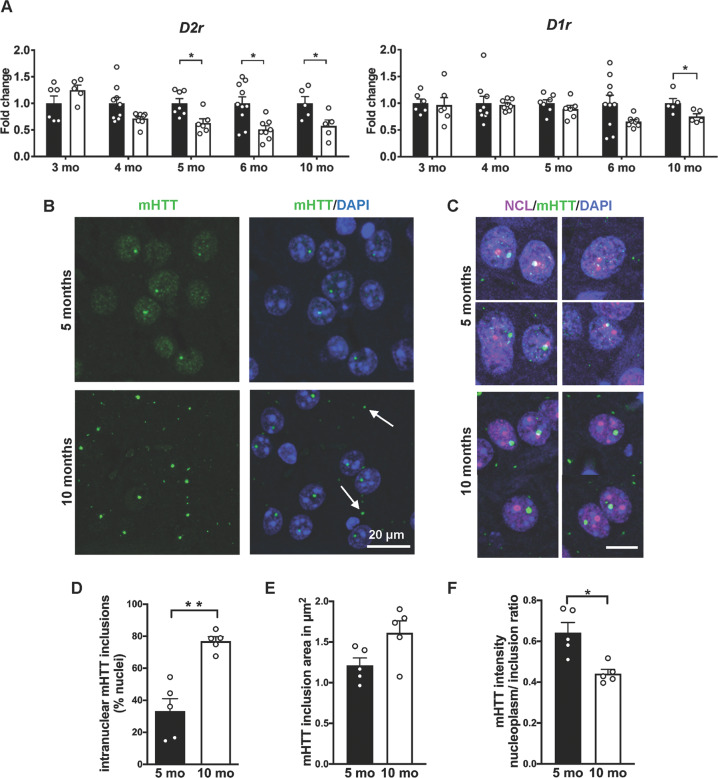
Transcriptional and cellular-stress surveillance deficits are hallmarks of Huntington's disease (HD), a fatal autosomal-dominant neurodegenerative disorder caused by a pathological expansion of CAG repeats in the Huntingtin (HTT) gene. The nucleolus, a dynamic nuclear biomolecular condensate and the site of ribosomal RNA (rRNA) transcription, is implicated in the cellular stress response and in protein quality control. While the exact pathomechanisms of HD are still unclear, the impact of nucleolar dysfunction on HD pathophysiology in vivo remains elusive. Here we identified aberrant maturation of rRNA and decreased translational rate in association with human mutant Huntingtin (mHTT) expression. The protein nucleophosmin 1 (NPM1), important for nucleolar integrity and rRNA maturation, loses its prominent nucleolar localization. Genetic disruption of nucleolar integrity in vulnerable striatal neurons of the R6/2 HD mouse model decreases the distribution of mHTT in a disperse state in the nucleus, exacerbating motor deficits. We confirmed NPM1 delocalization in the gradually progressing zQ175 knock-in HD mouse model: in the striatum at a presymptomatic stage and in the skeletal muscle at an early symptomatic stage. In Huntington's patient skeletal muscle biopsies, we found a selective redistribution of NPM1, similar to that in the zQ175 model. Taken together, our study demonstrates that nucleolar integrity regulates the formation of mHTT inclusions in vivo, and identifies NPM1 as a novel, readily detectable peripheral histopathological marker of HD progression.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Parlato R, Bierhoff H. Role of nucleolar dysfunction in neurodegenerative disorders: a game of genes? AIMS Mol Sci. 2015;2:211–24.
-
- Herrmann D, Parlato R. C9orf72-associated neurodegeneration in ALS-FTD: breaking new ground in ribosomal RNA and nucleolar dysfunction. Cell Tissue Res. 2018;373:351–60.. - PubMed
-
- Amer-Sarsour F, Ashkenazi A. The Nucleolus as a Proteostasis Regulator. Trends Cell Biol. 2019;29:849–51.. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
