

Randomized trial of neoadjuvant vaccination with tumor-cell lysate induces T cell response in low-grade gliomas
- PMID: 34882581
- PMCID: PMC8803342
- DOI: 10.1172/JCI151239
Randomized trial of neoadjuvant vaccination with tumor-cell lysate induces T cell response in low-grade gliomas
Abstract
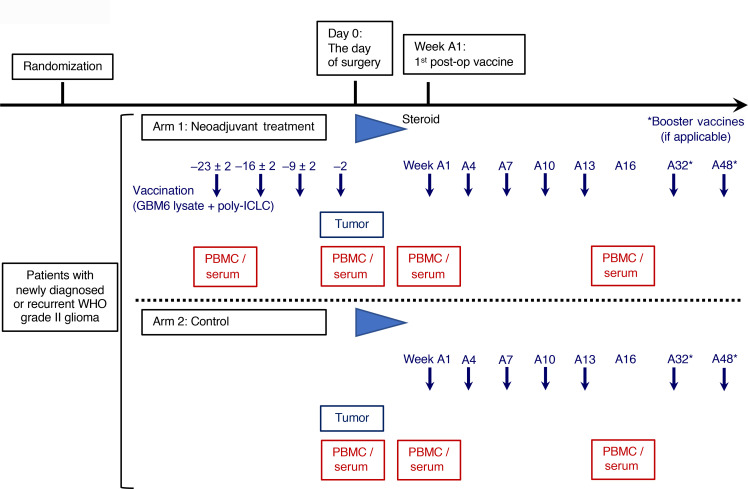
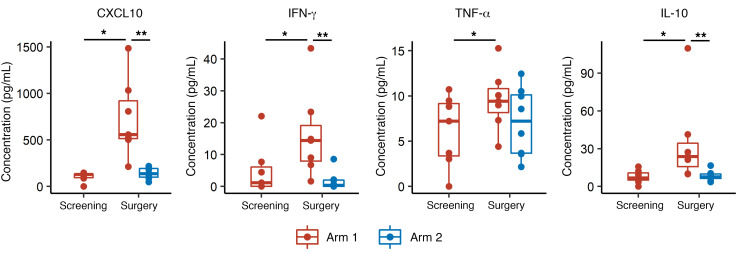
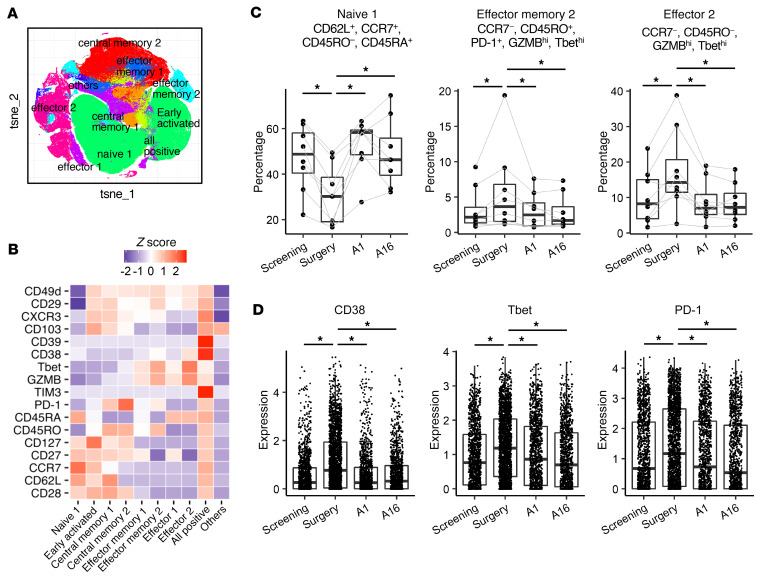
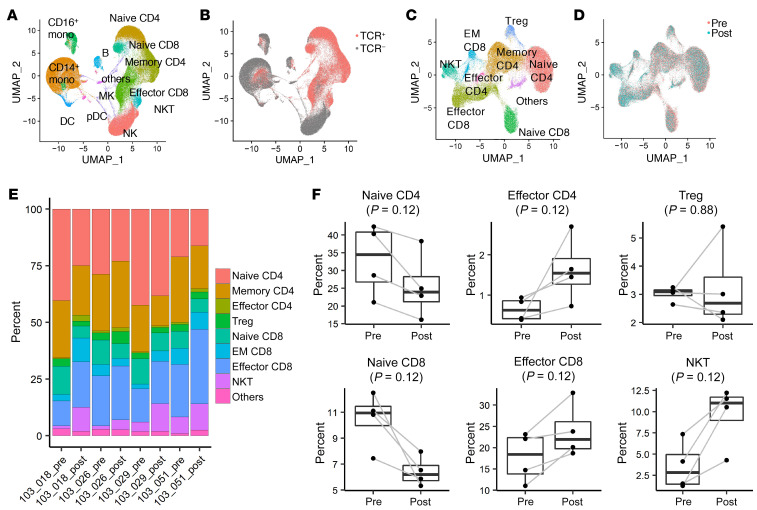
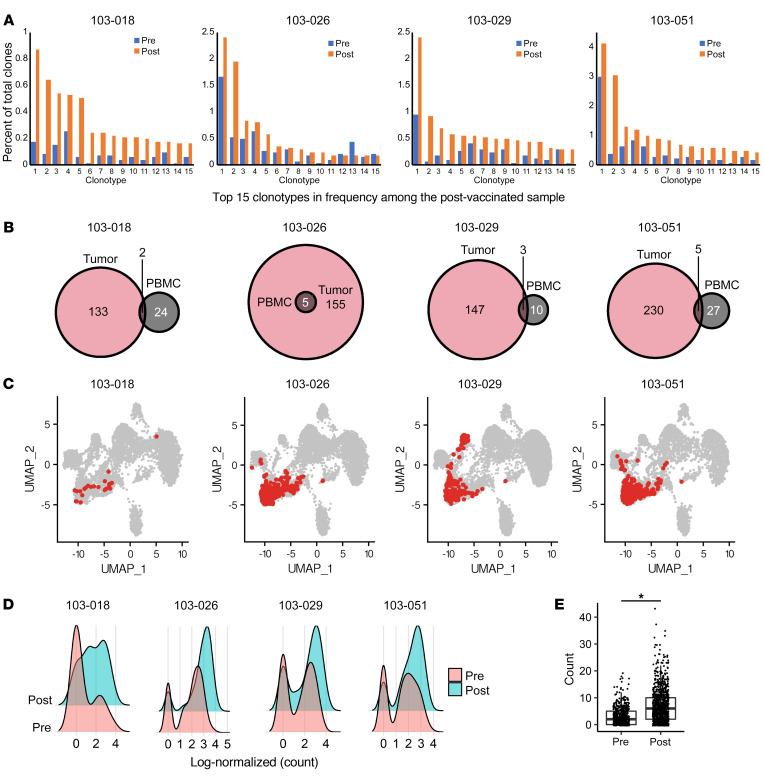
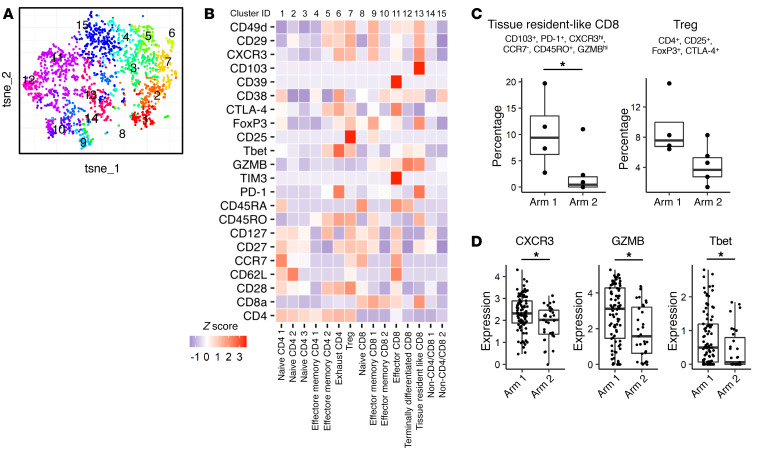
BACKGROUNDLong-term prognosis of WHO grade II low-grade gliomas (LGGs) is poor, with a high risk of recurrence and malignant transformation into high-grade gliomas. Given the relatively intact immune system of patients with LGGs and the slow tumor growth rate, vaccines are an attractive treatment strategy.METHODSWe conducted a pilot study to evaluate the safety and immunological effects of vaccination with GBM6-AD, lysate of an allogeneic glioblastoma stem cell line, with poly-ICLC in patients with LGGs. Patients were randomized to receive the vaccines before surgery (arm 1) or not (arm 2) and all patients received adjuvant vaccines. Coprimary outcomes were to evaluate safety and immune response in the tumor.RESULTSA total of 17 eligible patients were enrolled - 9 in arm 1 and 8 in arm 2. This regimen was well tolerated with no regimen-limiting toxicity. Neoadjuvant vaccination induced upregulation of type-1 cytokines and chemokines and increased activated CD8+ T cells in peripheral blood. Single-cell RNA/T cell receptor sequencing detected CD8+ T cell clones that expanded with effector phenotype and migrated into the tumor microenvironment (TME) in response to neoadjuvant vaccination. Mass cytometric analyses detected increased tissue resident-like CD8+ T cells with effector memory phenotype in the TME after the neoadjuvant vaccination.CONCLUSIONThe regimen induced effector CD8+ T cell response in peripheral blood and enabled vaccine-reactive CD8+ T cells to migrate into the TME. Further refinements of the regimen may have to be integrated into future strategies.TRIAL REGISTRATIONClinicalTrials.gov NCT02549833.FUNDINGNIH (1R35NS105068, 1R21CA233856), Dabbiere Foundation, Parker Institute for Cancer Immunotherapy, and Daiichi Sankyo Foundation of Life Science.
Keywords: Brain cancer; Cancer immunotherapy; Oncology; T cells; Vaccines.
Figures
References
-
- Brown PD, et al. Clinical Radiation Oncology. Churchill-Livingstone; 2006:493–514.
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
