

A new self-attenuated therapeutic influenza vaccine that uses host cell-restricted attenuation by artificial microRNAs
- PMID: 34883209
- PMCID: PMC8871448
- DOI: 10.1016/j.ijpharm.2021.121325
A new self-attenuated therapeutic influenza vaccine that uses host cell-restricted attenuation by artificial microRNAs
Abstract
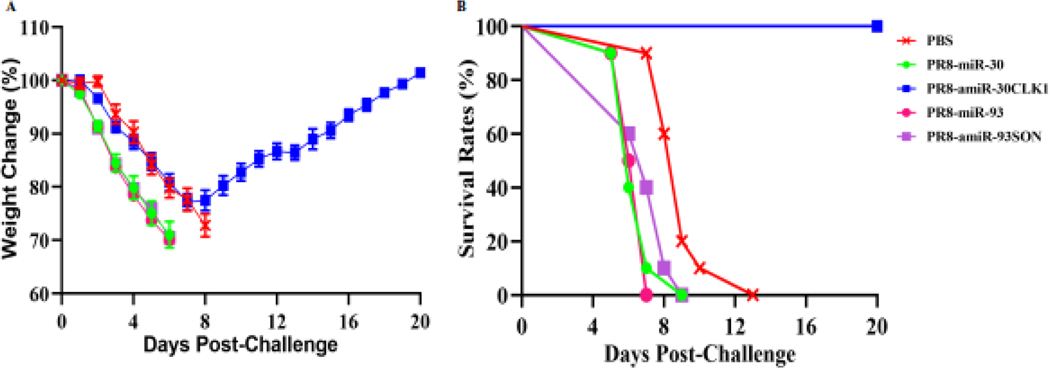
New strategies are urgently needed for developing vaccines and/or anti-viral drugs against influenza viruses, because antigenic shift and drift inevitably occurs in circulating strains each year, and new strains resistant to anti-viral drugs have recently emerged. In our study, we designed and incorporated artificial microRNAs (amiRNAs) into the NA segment of rescued influenza viruses to separately target two host genes, Cdc2-like kinase 1 (CLK1) and SON DNA binding protein (SON), which were found to play an essential role in virus replication. Mouse epithelial fibroblast (MEF) or human lung carcinoma A549 cells infected with engineered influenza PR8 viruses expressing amiR-30CLK1 (PR8-amiR-30CLK1) or amiR-93SON (PR8-amiR-93SON) had reduced expression of host proteins CLK1 and SON, respectively. All engineered influenza viruses functioned as attenuated vaccines, induced significantly higher antibody responses, and provided greater protective efficacy. In addition, they were found to be safe, based on the mouse weight changes and clinical signs observed. In contrast to the engineered viruses targeting SON, mice treated with engineered viruses targeting CLK1 recovered from weight loss and survived lethal infection by 6 h after lethal-dose PR8 infection, suggesting that our PR8-amiR-30CLK1 self-attenuated influenza virus (SAIV) could be used as a new therapeutic influenza vaccine.
Keywords: Artificial microRNA; Cdc2-like kinase 1; Intranasal delivery; Live attenuated influenza vaccine; SON DNA-binding protein; Therapeutic vaccine.
Copyright © 2021 Elsevier B.V. All rights reserved.
Figures





References
-
- Lozano R, Naghavi M, Foreman K, Lim S, Shibuya K, Aboyans V, Abraham J, Adair T, Aggarwal R, Ahn SY, Alvarado M, Anderson HR, Anderson LM, Andrews KG, Atkinson C, Baddour LM, Barker-Collo S, Bartels DH, Bell ML, Benjamin EJ, Bennett D, Bhalla K, Bikbov B, Bin Abdulhak A, Birbeck G, Blyth F, Bolliger I, Boufous S, Bucello C, Burch M, Burney P, Carapetis J, Chen H, Chou D, Chugh SS, Coffeng LE, Colan SD, Colquhoun S, Colson KE, Condon J, Connor MD, Cooper LT, Corriere M, Cortinovis M, de Vaccaro KC, Couser W, Cowie BC, Criqui MH, Cross M, Dabhadkar KC, Dahodwala N, De Leo D, Degenhardt L, Delossantos A, Denenberg J, Des Jarlais DC, Dharmaratne SD, Dorsey ER, Driscoll T, Duber H, Ebel B, Erwin PJ, Espindola P, Ezzati M, Feigin V, Flaxman AD, Forouzanfar MH, Fowkes FG, Franklin R, Fransen M, Freeman MK, Gabriel SE, Gakidou E, Gaspari F, Gillum RF, Gonzalez-Medina D, Halasa YA, Haring D, Harrison JE, Havmoeller R, Hay RJ, Hoen B, Hotez PJ, Hoy D, Jacobsen KH, James SL, Jasrasaria R, Jayaraman S, Johns N, Karthikeyan G, Kassebaum N, Keren A, Khoo JP, Knowlton LM, Kobusingye O, Koranteng A, Krishnamurthi R, Lipnick M, Lipshultz SE, Ohno SL, Mabweijano J, MacIntyre MF, Mallinger L, March L, Marks GB, Marks R, Matsumori A, Matzopoulos R, Mayosi BM, McAnulty JH, McDermott MM, McGrath J, Mensah GA, Merriman TR, Michaud C, Miller M, Miller TR, Mock C, Mocumbi AO, Mokdad AA, Moran A, Mulholland K, Nair MN, Naldi L, Narayan KM, Nasseri K, Norman P, O’Donnell M, Omer SB, Ortblad K, Osborne R, Ozgediz D, Pahari B, Pandian JD, Rivero AP, Padilla RP, Perez-Ruiz F, Perico N, Phillips D, Pierce K, Pope CA 3rd, Porrini E, Pourmalek F, Raju M, Ranganathan D, Rehm JT, Rein DB, Remuzzi G, Rivara FP, Roberts T, De Leon FR, Rosenfeld LC, Rushton L, Sacco RL, Salomon JA, Sampson U, Sanman E, Schwebel DC, Segui-Gomez M, Shepard DS, Singh D, Singleton J, Sliwa K, Smith E, Steer A, Taylor JA, Thomas B, Tleyjeh IM, Towbin JA, Truelsen T, Undurraga EA, Venketasubramanian N, Vijayakumar L, Vos T, Wagner GR, Wang M, Wang W, Watt K, Weinstock MA, Weintraub R, Wilkinson JD, Woolf AD, Wulf S, Yeh PH, Yip P, Zabetian A, Zheng ZJ, Lopez AD, Murray CJ, AlMazroa MA, Memish ZA, Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: a systematic analysis for the Global Burden of Disease Study 2010, Lancet, 380 (2012) 2095–2128. - PMC - PubMed
-
- Rudolf M, Poppel M, Frohlich A, Breithaupt A, Teifke J, Blohm U, Mettenleiter T, Beer M, Harder T, Longitudinal 2 years field study of conventional vaccination against highly pathogenic avian influenza H5N1 in layer hens, Vaccine, 28 (2010) 6832–6840. - PubMed
-
- Basha S, Hazenfeld S, Brady RC, Subbramanian RA, Comparison of antibody and T-cell responses elicited by licensed inactivated- and live-attenuated influenza vaccines against H3N2 hemagglutinin, Hum Immunol, 72 (2011) 463–469. - PubMed
-
- Hoft DF, Babusis E, Worku S, Spencer CT, Lottenbach K, Truscott SM, Abate G, Sakala IG, Edwards KM, Creech CB, Gerber MA, Bernstein DI, Newman F, Graham I, Anderson EL, Belshe RB, Live and inactivated influenza vaccines induce similar humoral responses, but only live vaccines induce diverse T-cell responses in young children, J Infect Dis, 204 (2011) 845–853. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
