

Canine DUXC: implications for DUX4 retrotransposition and preclinical models of FSHD
- PMID: 34888646
- PMCID: PMC9122657
- DOI: 10.1093/hmg/ddab352
Canine DUXC: implications for DUX4 retrotransposition and preclinical models of FSHD
Abstract
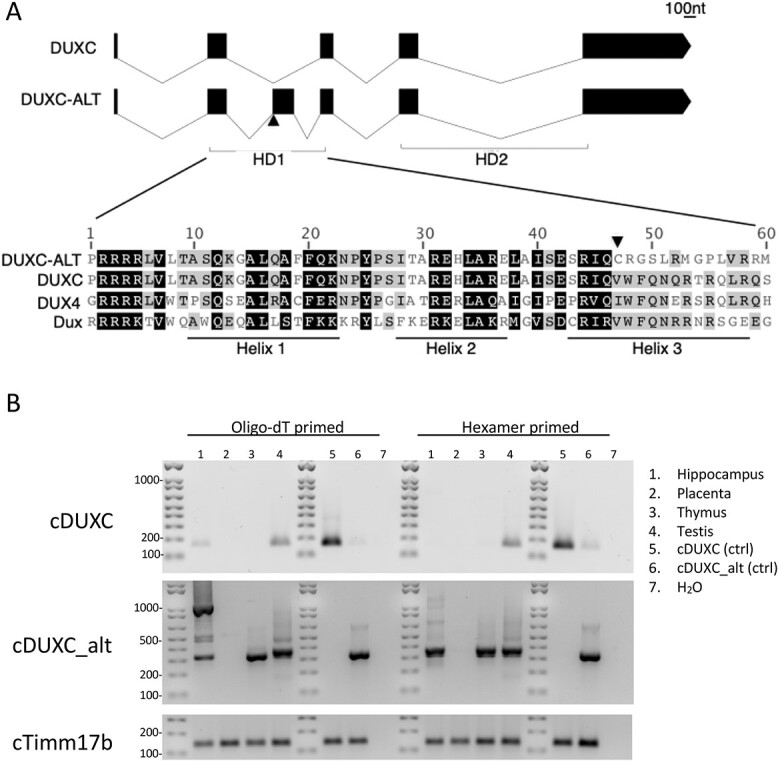
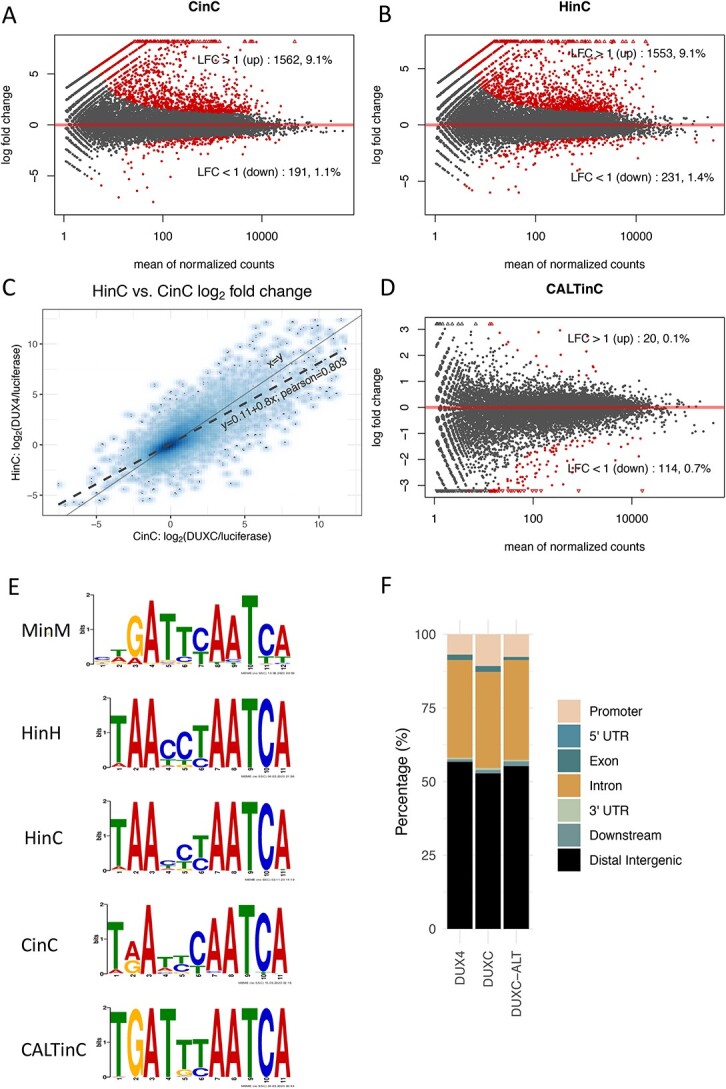
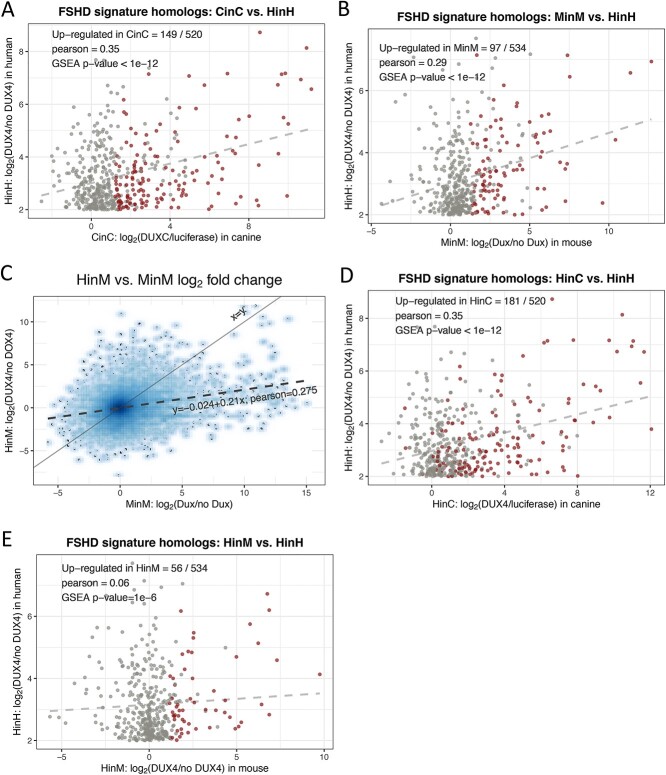
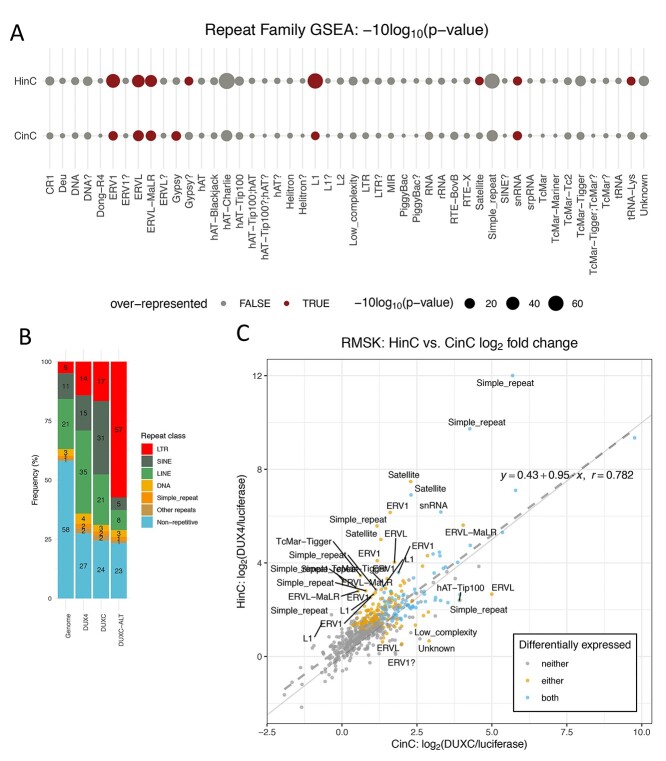
Mis-expression of DUX4 in skeletal muscle causes facioscapulohumeral muscular dystrophy (FSHD). Human DUX4 and mouse Dux are retrogenes derived from retrotransposition of the mRNA from the parental DUXC gene. Primates and rodents have lost the parental DUXC gene, and it is unknown whether DUXC had a similar role in driving an early pluripotent transcriptional program. Dogs and other Laurasiatherians have retained DUXC, providing an opportunity to determine the functional similarity to the retrotransposed DUX4 and Dux. Here, we identify the expression of two isoforms of DUXC mRNA in canine testis tissues: one encoding the canonical double homeodomain protein (DUXC), similar to DUX4/Dux, and a second that includes an in-frame alternative exon that disrupts the conserved amino acid sequence of the first homeodomain (DUXC-ALT). The expression of DUXC in canine cells induces a pluripotent program similar to DUX4 and Dux and induces the expression of a similar set of retrotransposons of the ERV/MaLR and LINE-1 families, as well as pericentromeric satellite repeats; whereas DUXC-ALT did not robustly activate gene expression in these assays. Important for preclinical models of FSHD, human DUX4 and canine DUXC show higher conservation of their homeodomains and corresponding binding motifs compared with the conservation between human DUX4 and mouse Dux, and human DUX4 activates a highly similar transcriptional program in canine cells. Together, these findings show that retrotransposition resulted in the loss of an alternatively spliced isoform and that DUXC containing mammals might be good candidates for certain preclinical models ofFSHD.
© The Author(s) 2021. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures
References
-
- Leidenroth, A., Clapp, J., Mitchell, L.M., Coneyworth, D., Dearden, F.L., Iannuzzi, L. and Hewitt, J.E. (2012) Evolution of DUX gene macrosatellites in placental mammals. Chromosoma, 121, 489–497. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
