

Dynamic regulation of N6,2'-O-dimethyladenosine (m6Am) in obesity
- PMID: 34893620
- PMCID: PMC8664860
- DOI: 10.1038/s41467-021-27421-2
Dynamic regulation of N6,2'-O-dimethyladenosine (m6Am) in obesity
Abstract
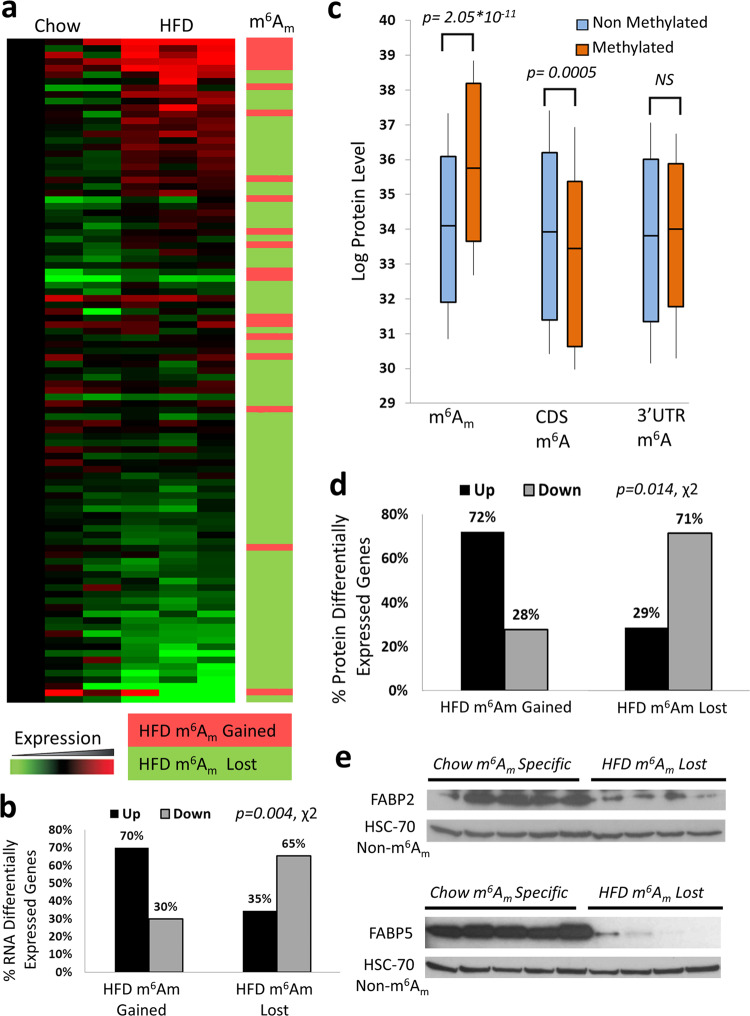
The prevalent m6Am mRNA cap modification was recently identified as a valid target for removal by the human obesity gene FTO along with the previously established m6A mRNA modification. However, the deposition and dynamics of m6Am in regulating obesity are unknown. Here, we investigate the liver m6A/m methylomes in mice fed on a high fat Western-diet and in ob/ob mice. We find that FTO levels are elevated in fat mice, and that genes which lost m6Am marking under obesity are overly downregulated, including the two fatty-acid-binding proteins FABP2, and FABP5. Furthermore, the cellular perturbation of FTO correspondingly affect protein levels of its targets. Notably, generally m6Am- but not m6A-methylated genes, are found to be highly enriched in metabolic processes. Finally, we deplete all m6A background via Mettl3 knockout, and unequivocally uncover the association of m6Am methylation with increased mRNA stability, translation efficiency, and higher protein expression. Together, these results strongly implicate a dynamic role for m6Am in obesity-related translation regulation.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Hess ME, et al. The fat mass and obesity-associated gene (Fto) regulates activity of the dopaminergic midbrain circuitry. Nat. Neurosci. 2013;16:1042–1048. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
