

Towards organoid culture without Matrigel
- PMID: 34893703
- PMCID: PMC8664924
- DOI: 10.1038/s42003-021-02910-8
Towards organoid culture without Matrigel
Abstract
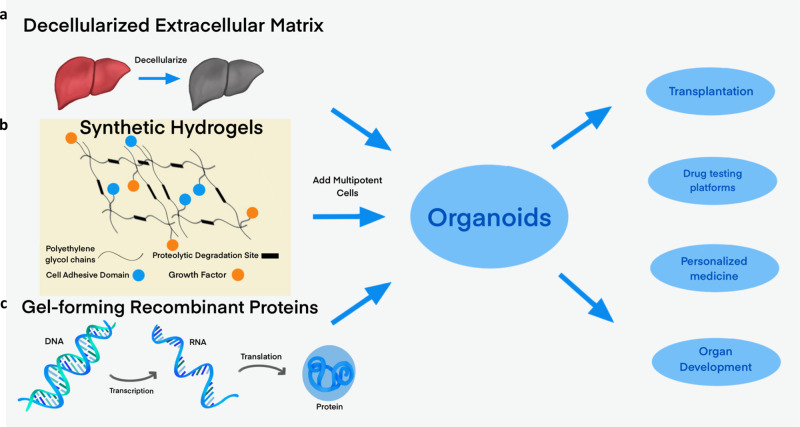
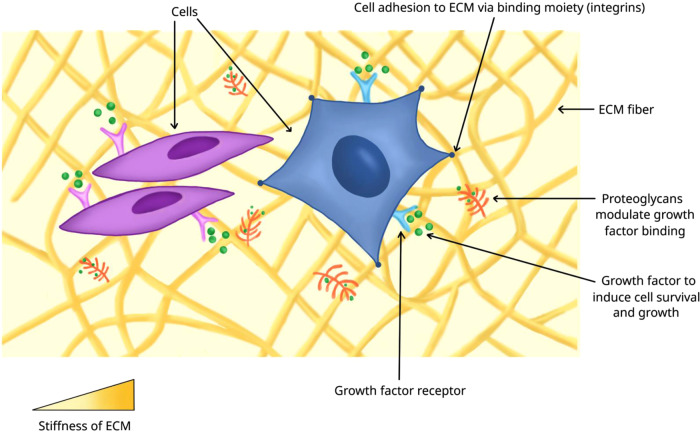
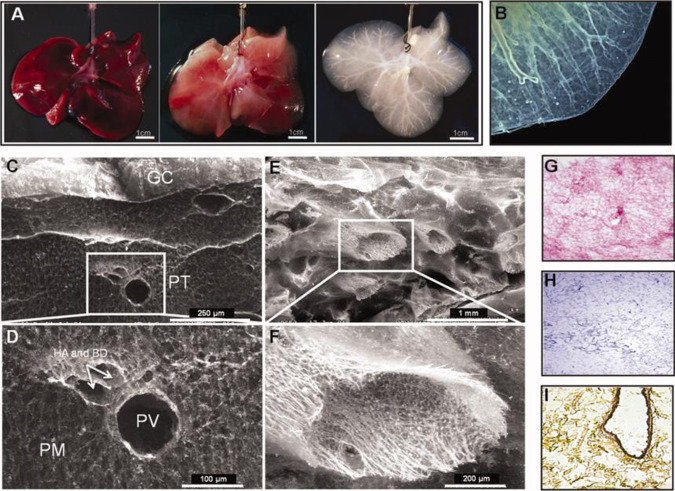
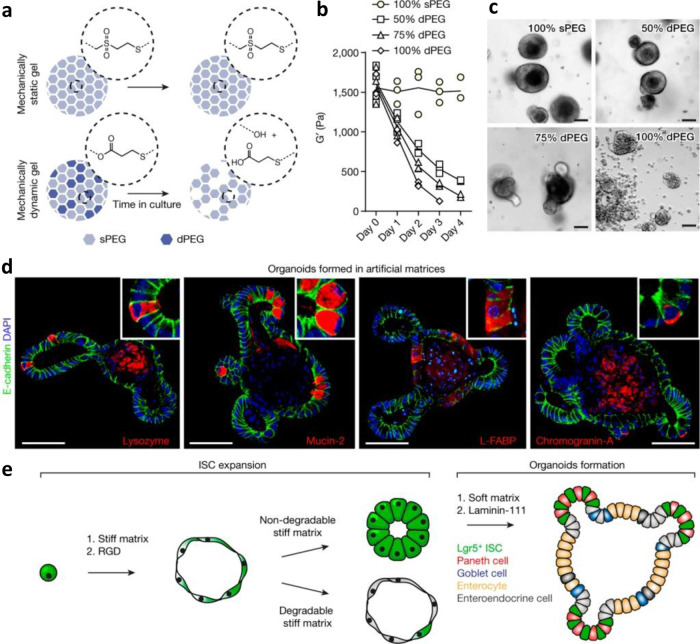
Organoids-cellular aggregates derived from stem or progenitor cells that recapitulate organ function in miniature-are of growing interest in developmental biology and medicine. Organoids have been developed for organs and tissues such as the liver, gut, brain, and pancreas; they are used as organ surrogates to study a wide range of questions in basic and developmental biology, genetic disorders, and therapies. However, many organoids reported to date have been cultured in Matrigel, which is prepared from the secretion of Engelbreth-Holm-Swarm mouse sarcoma cells; Matrigel is complex and poorly defined. This complexity makes it difficult to elucidate Matrigel-specific factors governing organoid development. In this review, we discuss promising Matrigel-free methods for the generation and maintenance of organoids that use decellularized extracellular matrix (ECM), synthetic hydrogels, or gel-forming recombinant proteins.
© 2021. This is a U.S. government work and not under copyright protection in the U.S.; foreign copyright protection may apply.
Conflict of interest statement
The authors declare no competing interests.
Figures
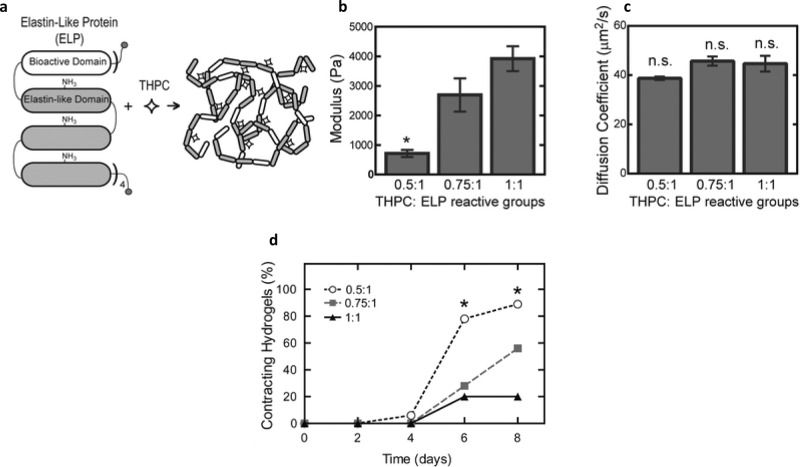
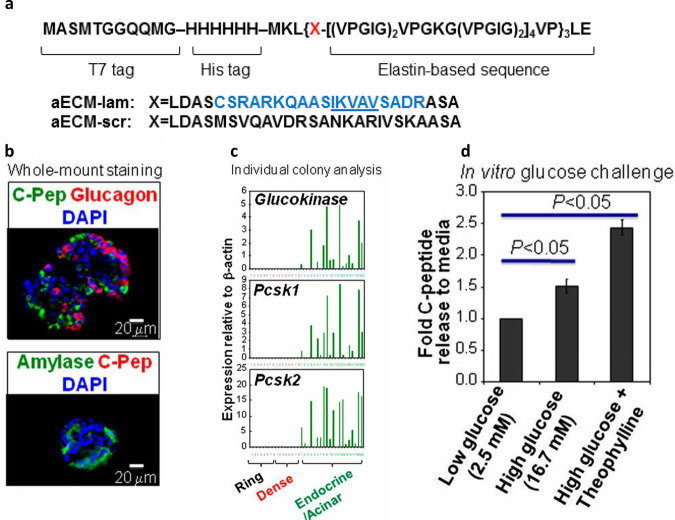
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
