

Whole-Genome Resequencing of Worldwide Wild and Domestic Sheep Elucidates Genetic Diversity, Introgression, and Agronomically Important Loci
- PMID: 34893856
- PMCID: PMC8826587
- DOI: 10.1093/molbev/msab353
Whole-Genome Resequencing of Worldwide Wild and Domestic Sheep Elucidates Genetic Diversity, Introgression, and Agronomically Important Loci
Abstract
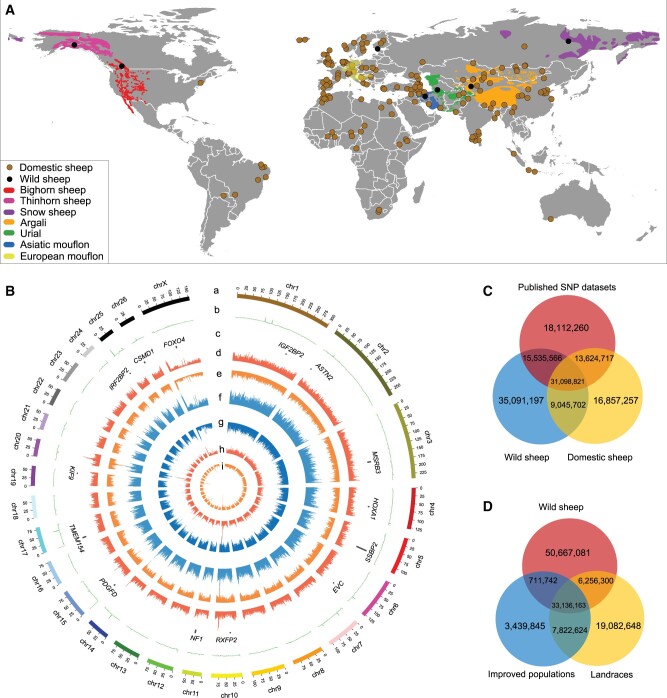
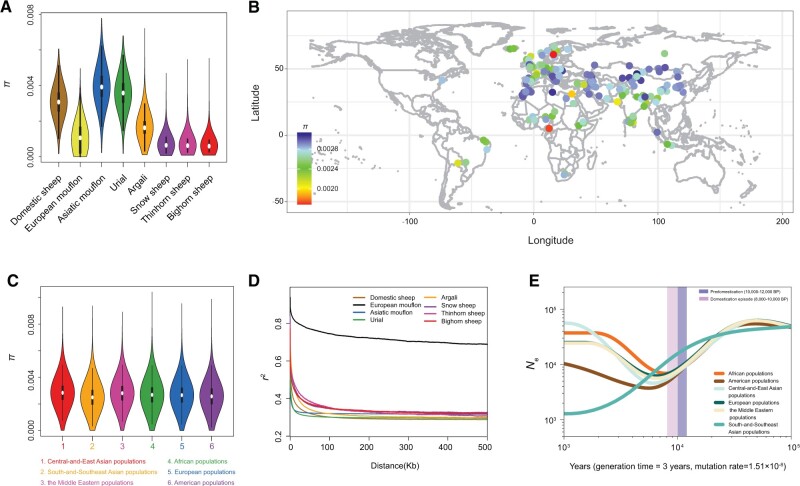
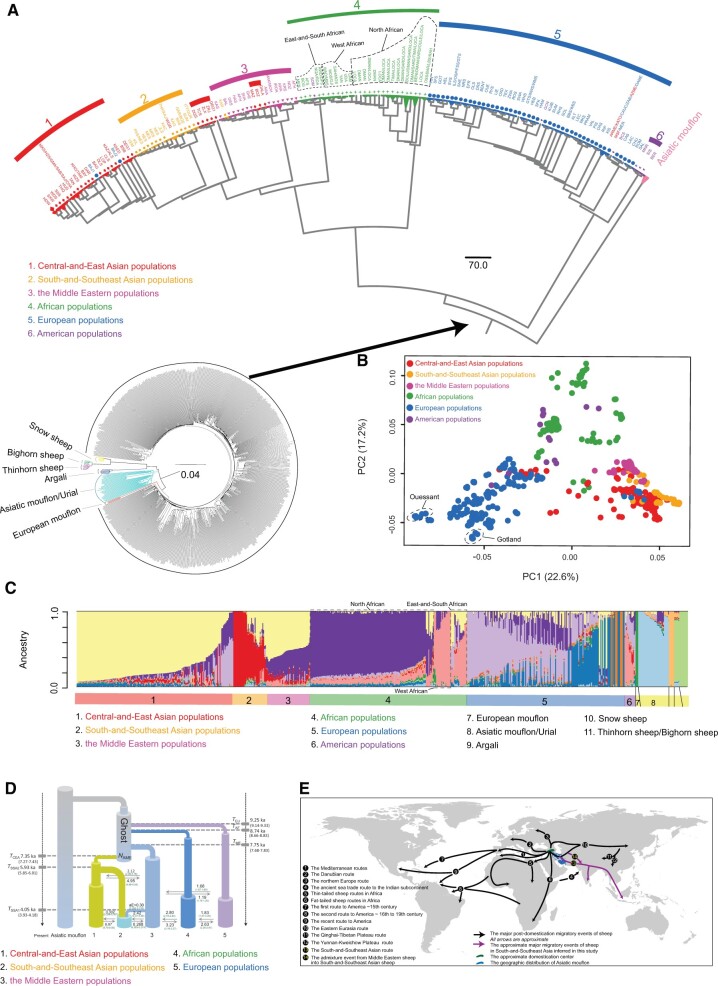
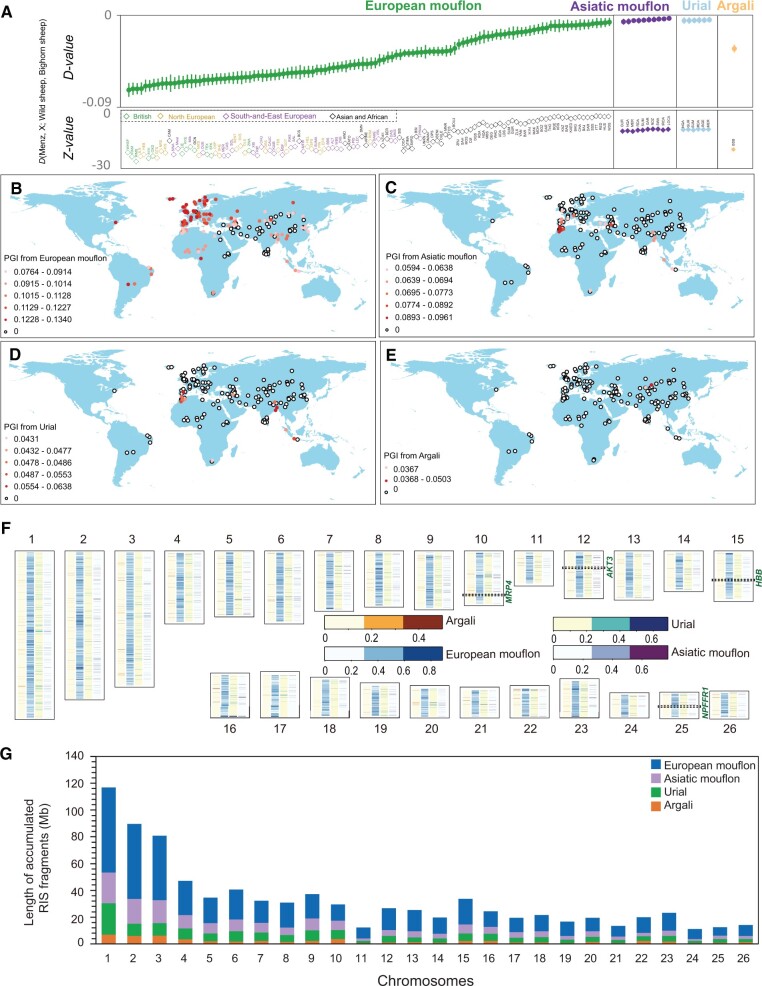
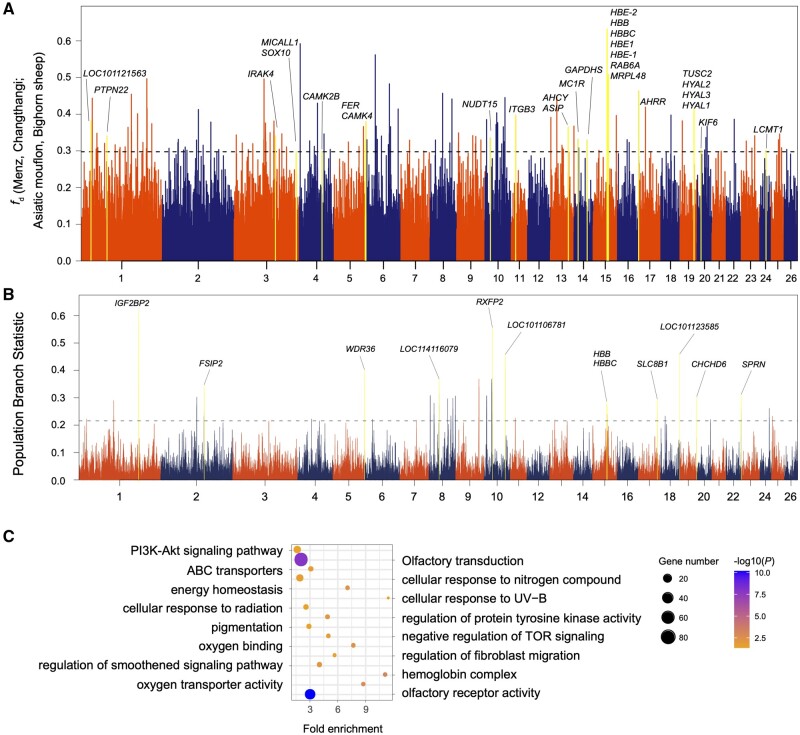
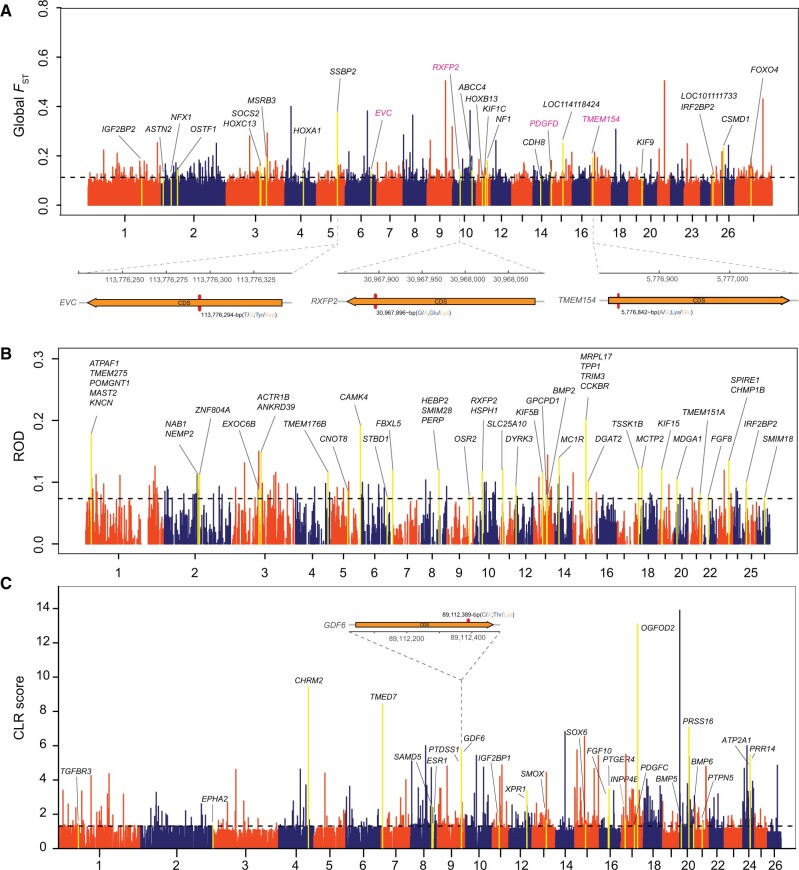
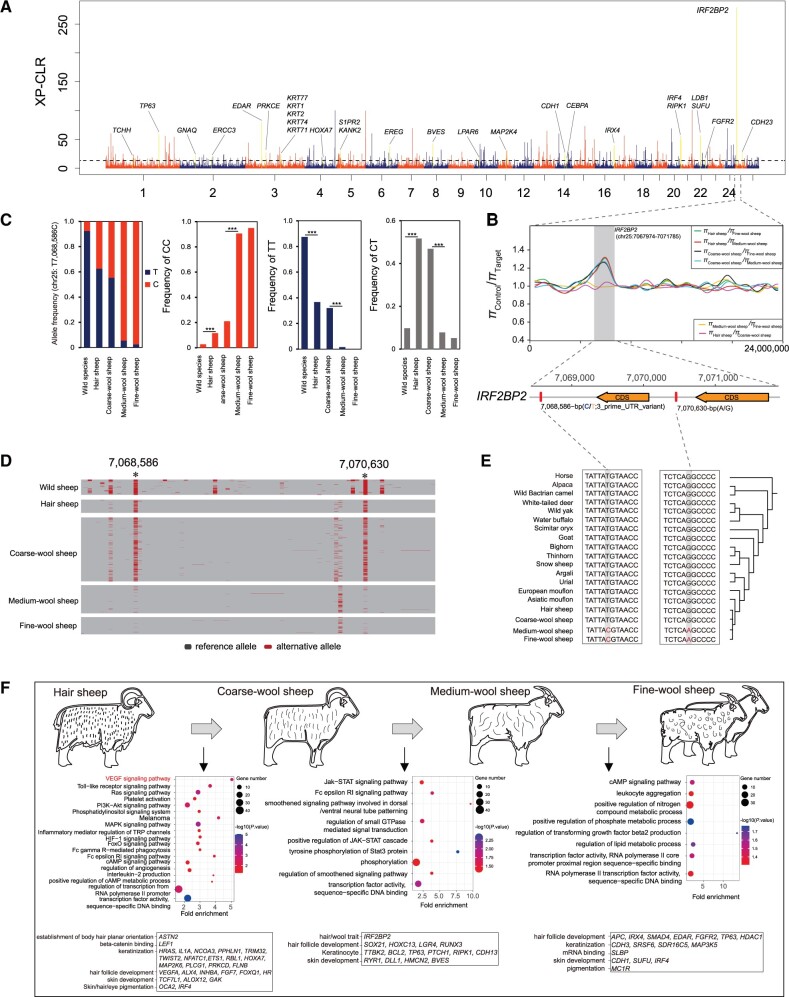
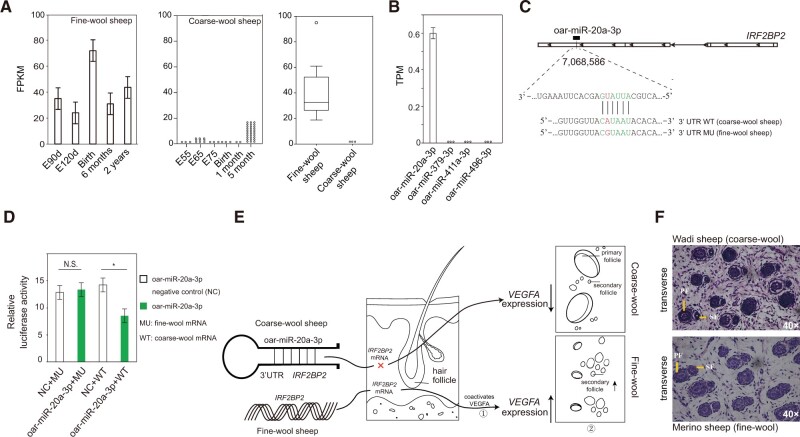
Domestic sheep and their wild relatives harbor substantial genetic variants that can form the backbone of molecular breeding, but their genome landscapes remain understudied. Here, we present a comprehensive genome resource for wild ovine species, landraces and improved breeds of domestic sheep, comprising high-coverage (∼16.10×) whole genomes of 810 samples from 7 wild species and 158 diverse domestic populations. We detected, in total, ∼121.2 million single nucleotide polymorphisms, ∼61 million of which are novel. Some display significant (P < 0.001) differences in frequency between wild and domestic species, or are private to continent-wide or individual sheep populations. Retained or introgressed wild gene variants in domestic populations have contributed to local adaptation, such as the variation in the HBB associated with plateau adaptation. We identified novel and previously reported targets of selection on morphological and agronomic traits such as stature, horn, tail configuration, and wool fineness. We explored the genetic basis of wool fineness and unveiled a novel mutation (chr25: T7,068,586C) in the 3'-UTR of IRF2BP2 as plausible causal variant for fleece fiber diameter. We reconstructed prehistorical migrations from the Near Eastern domestication center to South-and-Southeast Asia and found two main waves of migrations across the Eurasian Steppe and the Iranian Plateau in the Early and Late Bronze Ages. Our findings refine our understanding of genome variation as shaped by continental migrations, introgression, adaptation, and selection of sheep.
Keywords: adaptive introgression; agronomic traits; genetic diversity; genetic selection; migration; whole-genome sequences.
© The Author(s) 2021. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
References
-
- Ai H, Fang X, Yang B, Huang Z, Chen H, Mao L, Zhang F, Zhang L, Cui L, He W, et al.2015. Adaptation and possible ancient interspecies introgression in pigs identified by whole-genome sequencing. Nat Genet. 47(3):217–225. - PubMed
-
- Aldersey JE, Sonstegard TS, Williams JL, Bottema CDK.. 2020. Understanding the effects of the bovine POLLED variants. Anim Genet. 51(2):166–176. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
