

Loss of lamin-B1 and defective nuclear morphology are hallmarks of astrocyte senescence in vitro and in the aging human hippocampus
- PMID: 34894056
- PMCID: PMC8761005
- DOI: 10.1111/acel.13521
Loss of lamin-B1 and defective nuclear morphology are hallmarks of astrocyte senescence in vitro and in the aging human hippocampus
Abstract
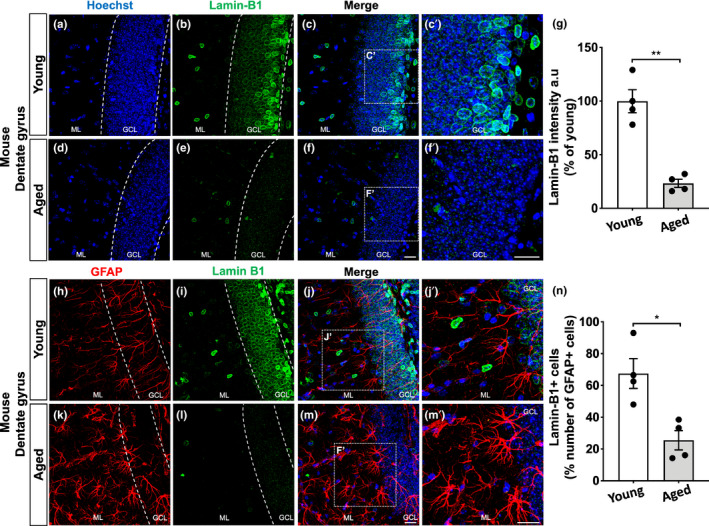
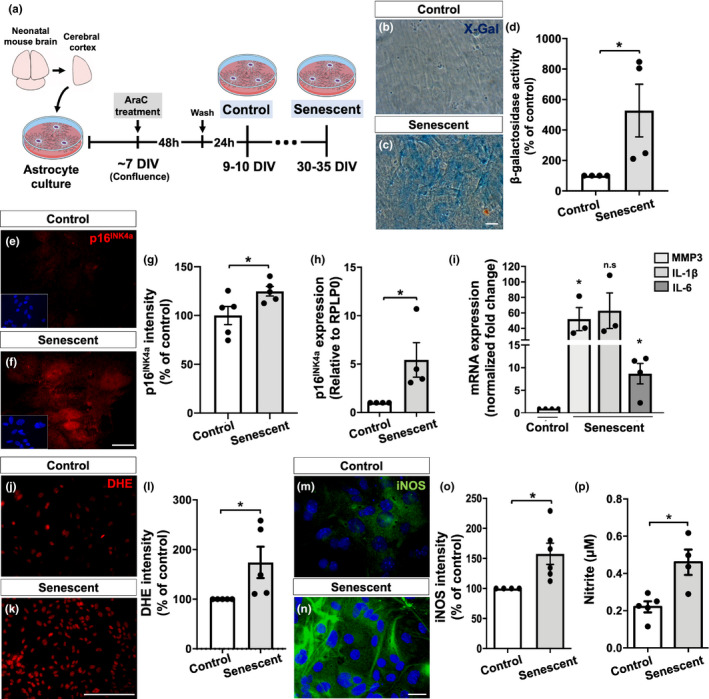
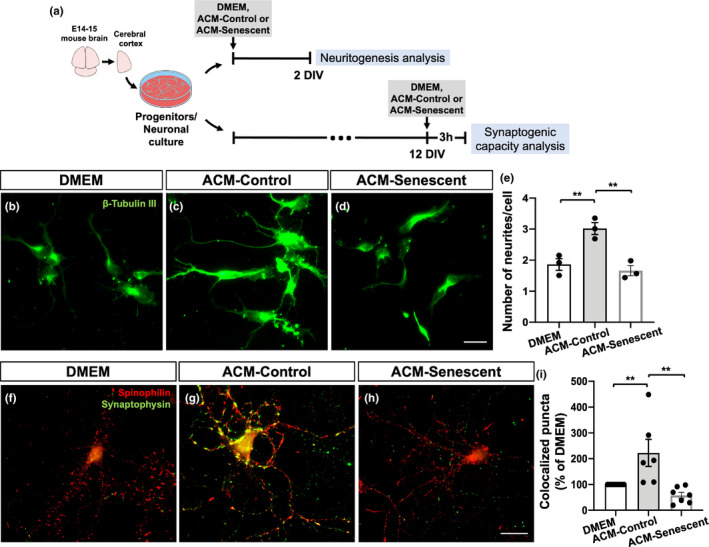
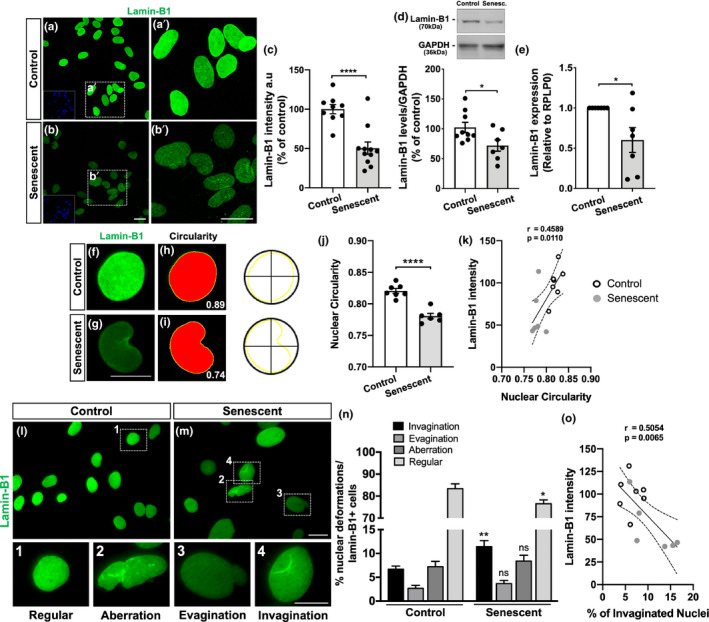
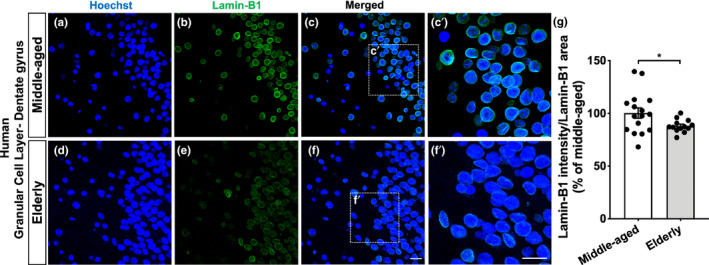
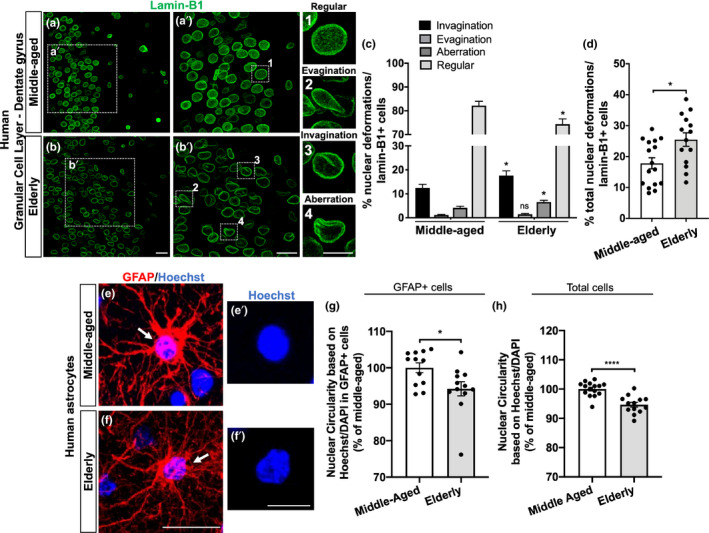
The increase in senescent cells in tissues, including the brain, is a general feature of normal aging and age-related pathologies. Senescent cells exhibit a specific phenotype, which includes an altered nuclear morphology and transcriptomic changes. Astrocytes undergo senescence in vitro and in age-associated neurodegenerative diseases, but little is known about whether this process also occurs in physiological aging, as well as its functional implication. Here, we investigated astrocyte senescence in vitro, in old mouse brains, and in post-mortem human brain tissue of elderly. We identified a significant loss of lamin-B1, a major component of the nuclear lamina, as a hallmark of senescent astrocytes. We showed a severe reduction of lamin-B1 in the dentate gyrus of aged mice, including in hippocampal astrocytes, and in the granular cell layer of the hippocampus of post-mortem human tissue from non-demented elderly. The lamin-B1 reduction was associated with nuclear deformations, represented by an increased incidence of invaginated nuclei and loss of nuclear circularity in senescent astrocytes in vitro and in the aging human hippocampus. We also found differences in lamin-B1 levels and astrocyte nuclear morphology between the granular cell layer and polymorphic layer in the elderly human hippocampus, suggesting an intra-regional-dependent aging response of human astrocytes. Moreover, we described senescence-associated impaired neuritogenic and synaptogenic capacity of mouse astrocytes. Our findings show that reduction of lamin-B1 is a conserved feature of hippocampal cells aging, including astrocytes, and shed light on significant defects in nuclear lamina structure which may contribute to astrocyte dysfunctions during aging.
Keywords: aging; astrocyte; human and mouse hippocampus; lamin-B1; senescence; synapse.
© 2021 The Authors. Aging Cell published by Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Barbar, L. , Jain, T. , Zimmer, M. , Kruglikov, I. , Sadick, J. S. , Wang, M. , Kalpana, K. , Rose, I. V. L. , Burstein, S. R. , Rusielewicz, T. , Nijsure, M. , Guttenplan, K. A. , di Domenico, A. , Croft, G. , Zhang, B. , Nobuta, H. , Hébert, J. M. , Liddelow, S. A. , & Fossati, V. (2020). CD49f is a novel marker of functional and reactive human iPSC‐derived astrocytes. Neuron, 107(3), 436–453.e12. 10.1016/j.neuron.2020.05.014 - DOI - PMC - PubMed
-
- Bedrosian, T. A. , Houtman, J. , Eguiguren, J. S. , Ghassemzadeh, S. , Rund, N. , Novaresi, N. M. , Hu, L. , Parylak, S. L. , Denli, A. M. , Randolph‐Moore, L. , Namba, T. , Gage, F. H. , & Toda, T. (2020). Lamin B1 decline underlies age‐related loss of adult hippocampal neurogenesis. EMBO Journal, 40(3), e105819. 10.15252/embj.2020105819 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
