

NADH inhibition of SIRT1 links energy state to transcription during time-restricted feeding
- PMID: 34903884
- PMCID: PMC8688143
- DOI: 10.1038/s42255-021-00498-1
NADH inhibition of SIRT1 links energy state to transcription during time-restricted feeding
Abstract
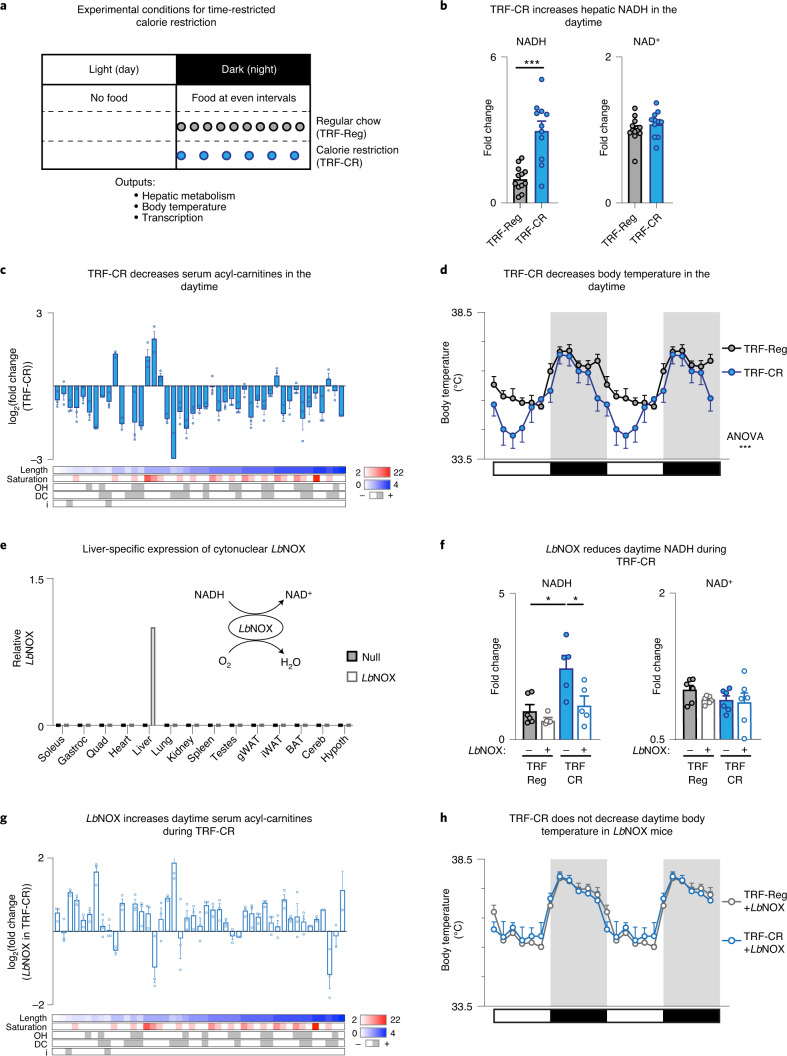
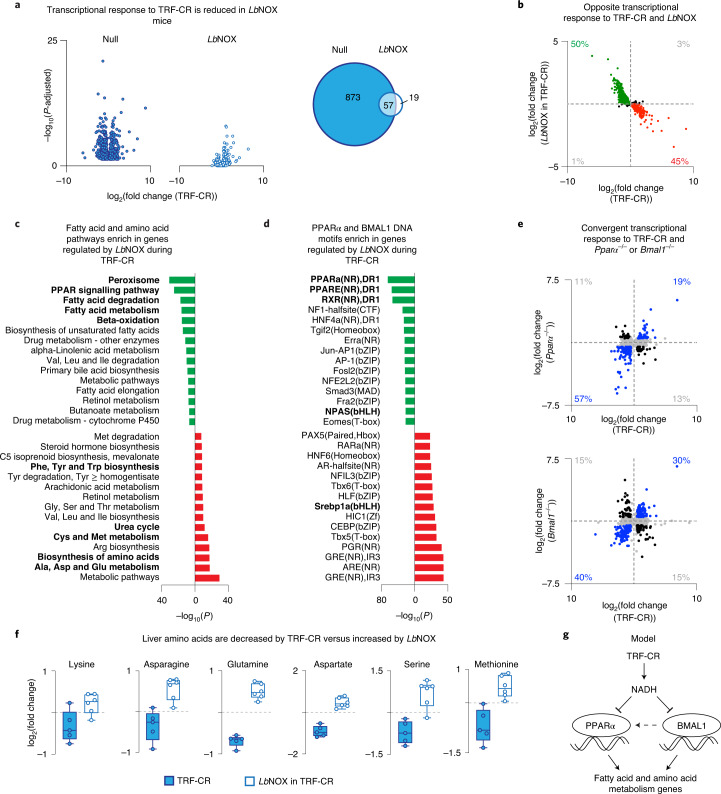
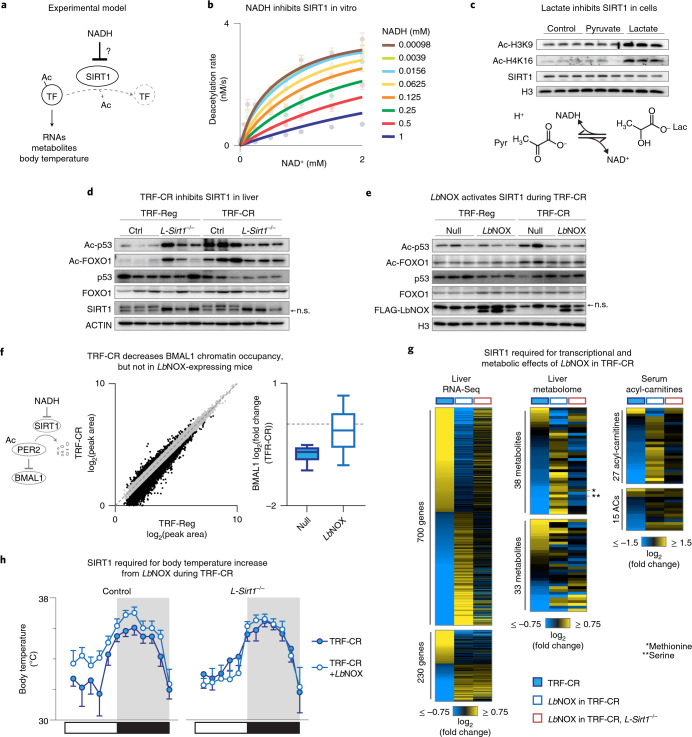
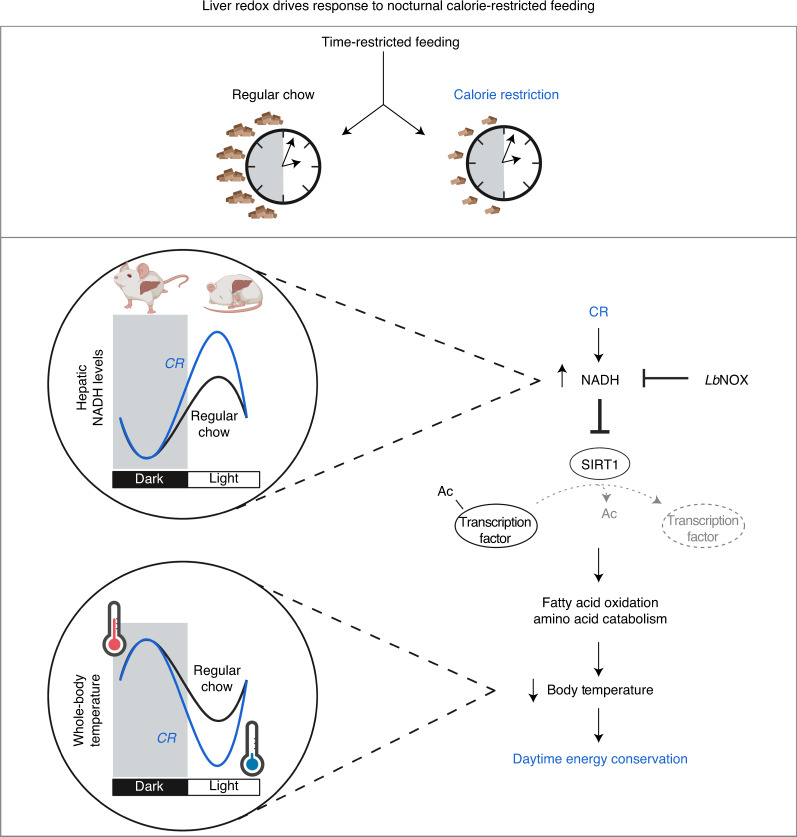
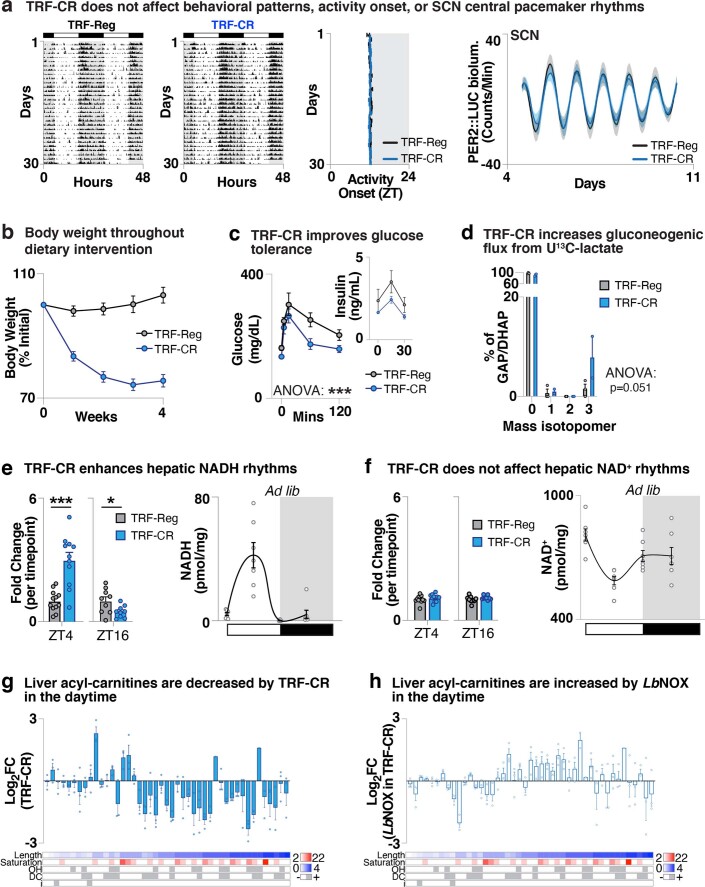
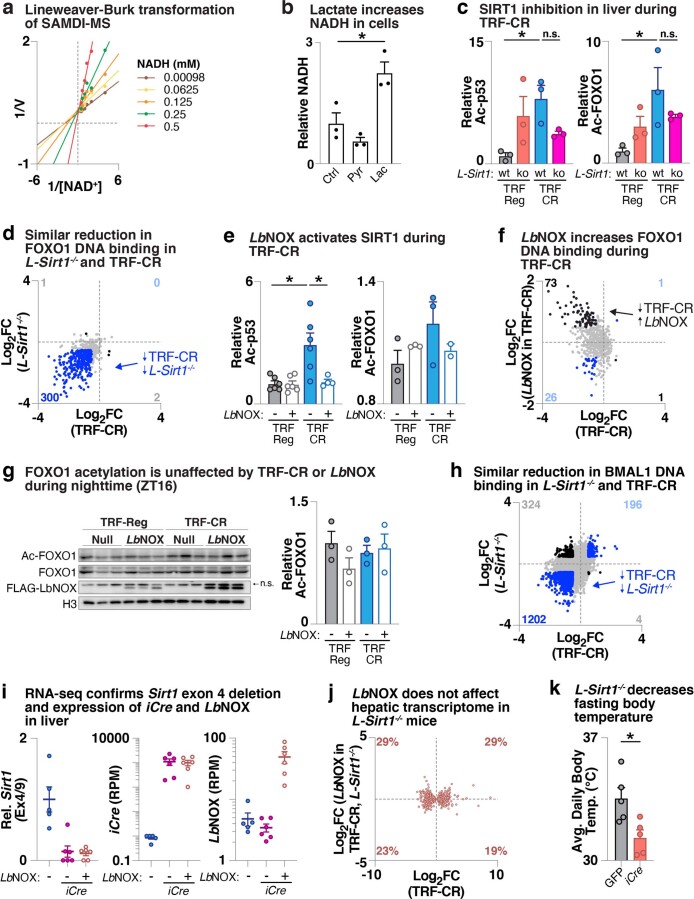
In mammals, circadian rhythms are entrained to the light cycle and drive daily oscillations in levels of NAD+, a cosubstrate of the class III histone deacetylase sirtuin 1 (SIRT1) that associates with clock transcription factors. Although NAD+ also participates in redox reactions, the extent to which NAD(H) couples nutrient state with circadian transcriptional cycles remains unknown. Here we show that nocturnal animals subjected to time-restricted feeding of a calorie-restricted diet (TRF-CR) only during night-time display reduced body temperature and elevated hepatic NADH during daytime. Genetic uncoupling of nutrient state from NADH redox state through transduction of the water-forming NADH oxidase from Lactobacillus brevis (LbNOX) increases daytime body temperature and blood and liver acyl-carnitines. LbNOX expression in TRF-CR mice induces oxidative gene networks controlled by brain and muscle Arnt-like protein 1 (BMAL1) and peroxisome proliferator-activated receptor alpha (PPARα) and suppresses amino acid catabolic pathways. Enzymatic analyses reveal that NADH inhibits SIRT1 in vitro, corresponding with reduced deacetylation of SIRT1 substrates during TRF-CR in vivo. Remarkably, Sirt1 liver nullizygous animals subjected to TRF-CR display persistent hypothermia even when NADH is oxidized by LbNOX. Our findings reveal that the hepatic NADH cycle links nutrient state to whole-body energetics through the rhythmic regulation of SIRT1.
© 2021. The Author(s).
Conflict of interest statement
The authors declare the following competing financial interest(s): M.M. is founder and chairman of SAMDI Tech Inc., which uses SAMDI-MS to perform high-throughput screening and assay chemistry to clients in the pharmaceutical industry. All other authors declare no competing financial interests.
Figures

Comment in
-
Delving into the effects of time-restricted feeding.Nat Rev Endocrinol. 2022 Mar;18(3):134. doi: 10.1038/s41574-021-00628-5. Nat Rev Endocrinol. 2022. PMID: 34949780 No abstract available.
References
-
- Bass J, Lazar MA. Circadian time signatures of fitness and disease. Science. 2016;354:994–999. - PubMed
-
- Lin S-J, et al. Calorie restriction extends Saccharomyces cerevisiae lifespan by increasing respiration. Nature. 2002;418:344–348. - PubMed
-
- Weindruch R, Walford RL, Fligiel S, Guthrie D. The retardation of aging in mice by dietary restriction: longevity, cancer, immunity and lifetime energy intake. J. Nutr. 1986;116:641–654. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK090625/DK/NIDDK NIH HHS/United States
- R03 DK116012/DK/NIDDK NIH HHS/United States
- P01 AG049665/AG/NIA NIH HHS/United States
- T32 GM008061/GM/NIGMS NIH HHS/United States
- R01 DK108987/DK/NIDDK NIH HHS/United States
- T32 HL007909/HL/NHLBI NIH HHS/United States
- R01 AG065988/AG/NIA NIH HHS/United States
- R35 CA197532/CA/NCI NIH HHS/United States
- P01 AG011412/AG/NIA NIH HHS/United States
- U54 CA199091/CA/NCI NIH HHS/United States
- R01 HD089552/HD/NICHD NIH HHS/United States
- R01 DK100814/DK/NIDDK NIH HHS/United States
- R01 DK113011/DK/NIDDK NIH HHS/United States
- F32 DK122675/DK/NIDDK NIH HHS/United States
- K01 DK105137/DK/NIDDK NIH HHS/United States
- R01 DK127800/DK/NIDDK NIH HHS/United States
- R01 DK050203/DK/NIDDK NIH HHS/United States
- P30 DK124723/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
