

The Hippo Pathway Effectors YAP and TAZ Regulate LH Release by Pituitary Gonadotrope Cells in Mice
- PMID: 34905605
- PMCID: PMC8670590
- DOI: 10.1210/endocr/bqab238
The Hippo Pathway Effectors YAP and TAZ Regulate LH Release by Pituitary Gonadotrope Cells in Mice
Abstract
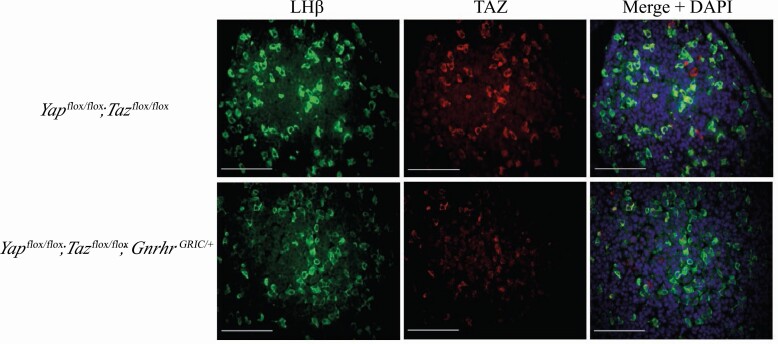
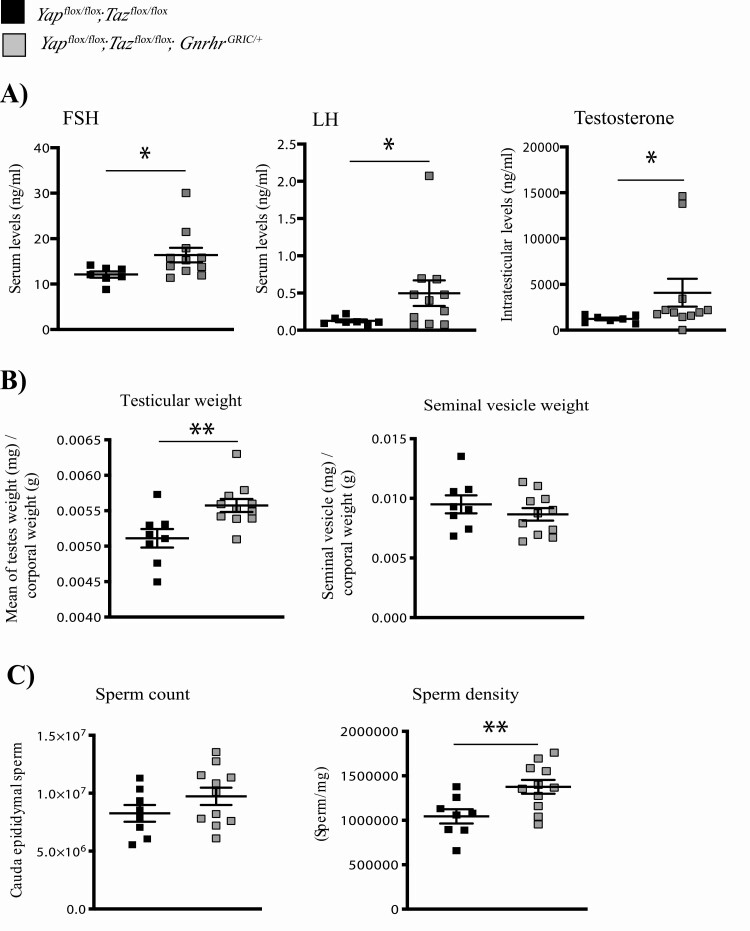
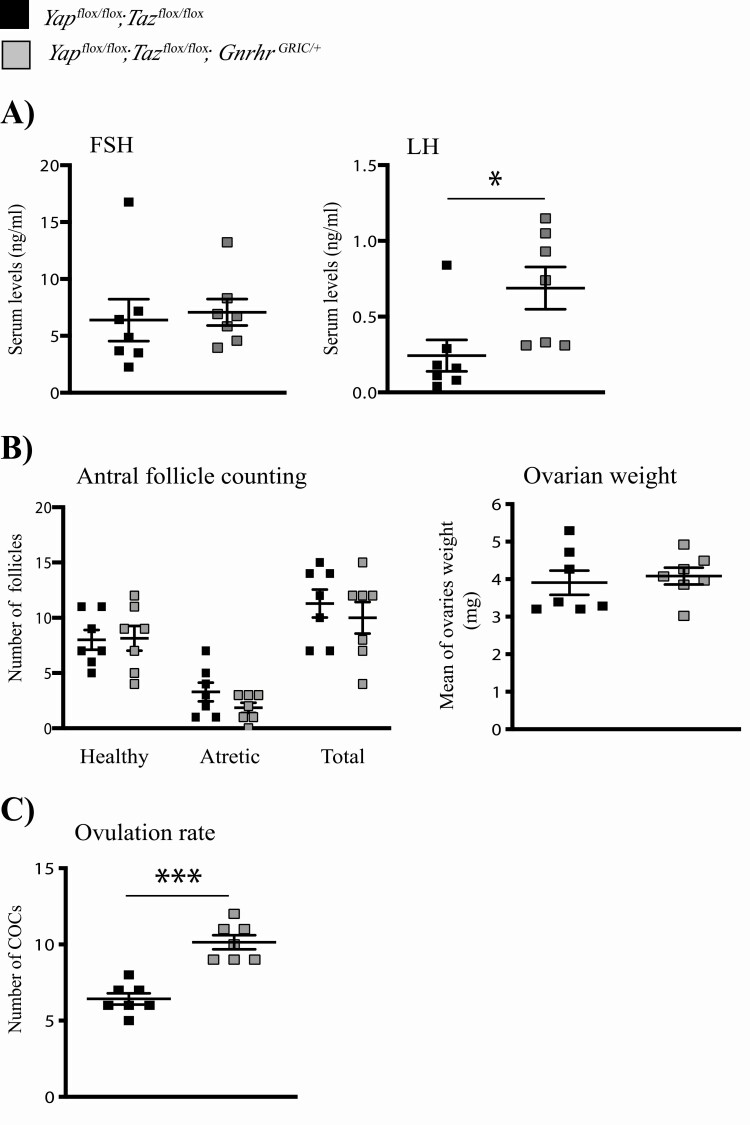
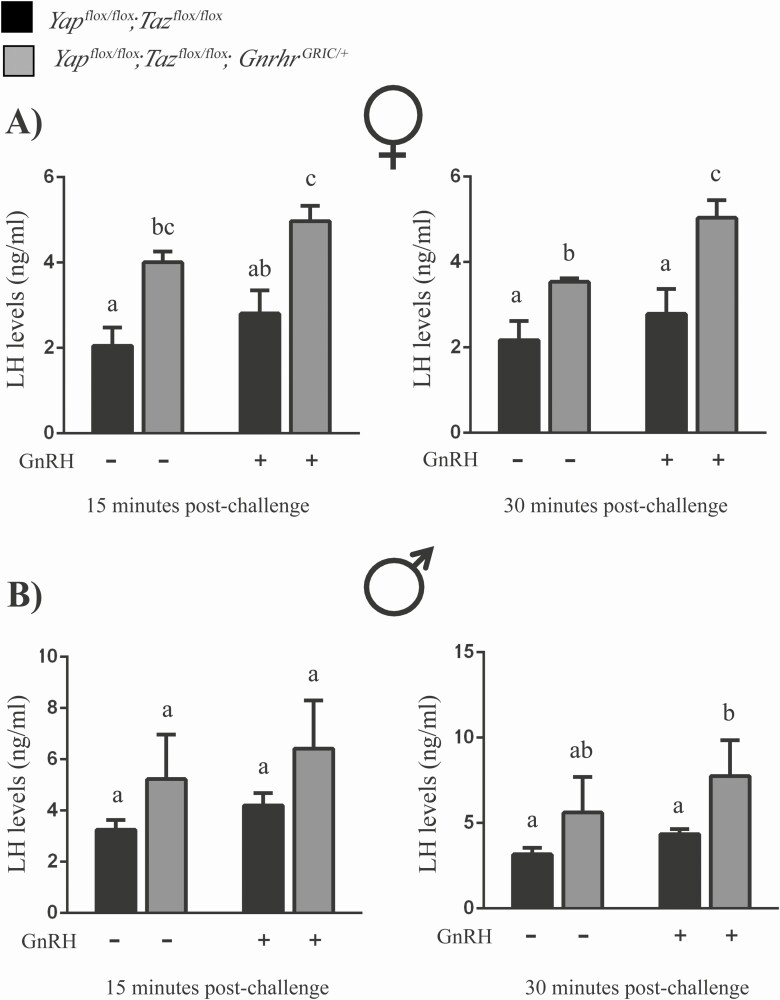
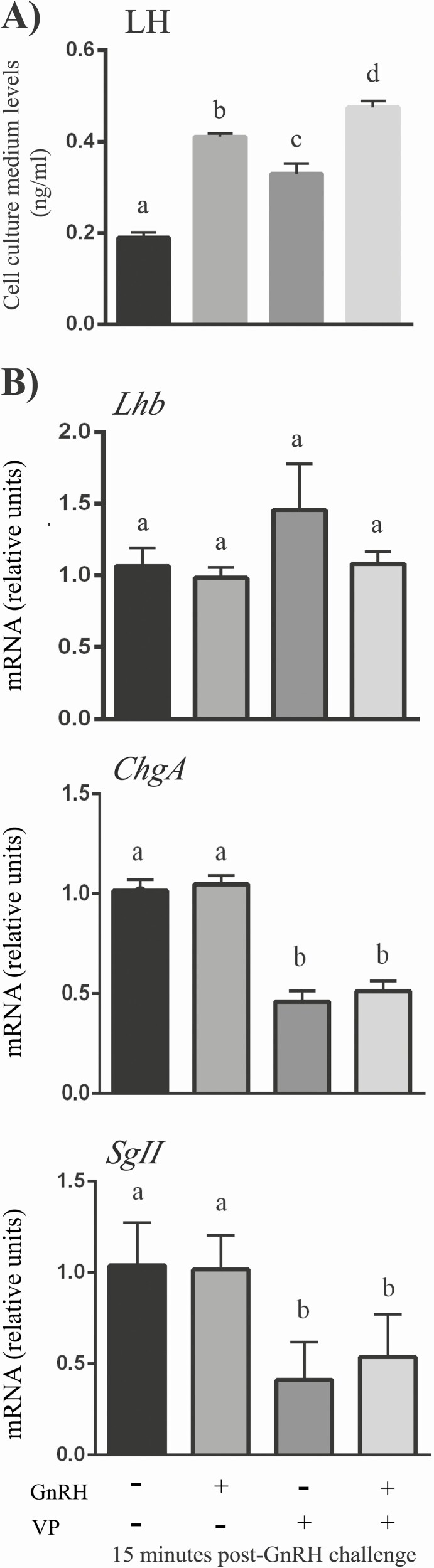
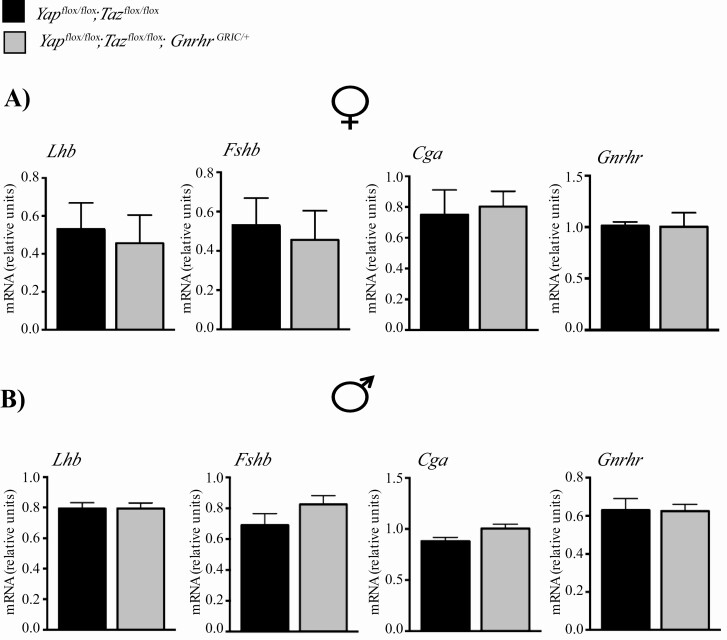
The Hippo transcriptional coactivators YAP and TAZ exert critical roles in morphogenesis, organ size determination and tumorigenesis in many tissues. Although Hippo kinase cascade activity was recently reported in the anterior pituitary gland in mice, the role of the Hippo effectors in regulating gonadotropin production remains unknown. The objective of this study was therefore to characterize the roles of YAP and TAZ in gonadotropin synthesis and secretion. Using a conditional gene targeting approach (cKO), we found that gonadotrope-specific inactivation of Yap and Taz resulted in increased circulating levels of follicle-stimulating hormone (FSH) and luteinizing hormone (LH) in adult male mice, along with increased testosterone levels and testis weight. Female cKO mice had increased circulating LH (but not FSH) levels, which were associated with a hyperfertility phenotype characterized by higher ovulation rates and larger litter sizes. Unexpectedly, the loss of YAP/TAZ did not appear to affect the expression of gonadotropin subunit genes, yet both basal and GnRH-induced LH secretion were increased in cultured pituitary cells from cKO mice. Likewise, pharmacologic inhibition of YAP binding to the TEAD family of transcription factors increased both basal and GnRH-induced LH secretion in LβT2 gonadotrope-like cells in vitro without affecting Lhb expression. Conversely, mRNA levels of ChgA and SgII, which encode key secretory granule cargo proteins, were decreased following pharmacologic inhibition of YAP/TAZ, suggesting a mechanism whereby YAP/TAZ regulate the LH secretion machinery in gonadotrope cells. Together, these findings represent the first evidence that Hippo signaling may play a role in regulating pituitary LH secretion.
Keywords: KO mice; TAZ; YAP; fertility; gonadotropins.
© The Author(s) 2021. Published by Oxford University Press on behalf of the Endocrine Society. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
Similar articles
-
Deletion of Gαq/11 or Gαs Proteins in Gonadotropes Differentially Affects Gonadotropin Production and Secretion in Mice.Endocrinology. 2022 Feb 1;163(2):bqab247. doi: 10.1210/endocr/bqab247. Endocrinology. 2022. PMID: 34864945 Free PMC article.
-
NR5A2 regulates Lhb and Fshb transcription in gonadotrope-like cells in vitro, but is dispensable for gonadotropin synthesis and fertility in vivo.PLoS One. 2013;8(3):e59058. doi: 10.1371/journal.pone.0059058. Epub 2013 Mar 11. PLoS One. 2013. PMID: 23536856 Free PMC article.
-
GnRH Receptor Expression and Reproductive Function Depend on JUN in GnRH Receptor‒Expressing Cells.Endocrinology. 2018 Mar 1;159(3):1496-1510. doi: 10.1210/en.2017-00844. Endocrinology. 2018. PMID: 29409045 Free PMC article.
-
The differential secretion of FSH and LH: regulation through genes, feedback and packaging.Reprod Suppl. 2003;61:463-76. Reprod Suppl. 2003. PMID: 14635955 Review.
-
Discovery of new receptors regulating luteinizing hormone and follicle-stimulating hormone secretion by bovine gonadotrophs to explore a new paradigm for mechanisms regulating reproduction.J Reprod Dev. 2020 Aug 20;66(4):291-297. doi: 10.1262/jrd.2020-012. Epub 2020 Apr 6. J Reprod Dev. 2020. PMID: 32249236 Free PMC article. Review.
Cited by
-
Pituitary Tumorigenesis-Implications for Management.Medicina (Kaunas). 2023 Apr 21;59(4):812. doi: 10.3390/medicina59040812. Medicina (Kaunas). 2023. PMID: 37109772 Free PMC article. Review.
-
The dynamic expression of YAP is essential for the development of male germ cells derived from human embryonic stem cells.Sci Rep. 2024 Jul 8;14(1):15732. doi: 10.1038/s41598-024-66852-x. Sci Rep. 2024. PMID: 38977826 Free PMC article.
-
Hippo Signaling in the Ovary: Emerging Roles in Development, Fertility, and Disease.Endocr Rev. 2022 Nov 25;43(6):1074-1096. doi: 10.1210/endrev/bnac013. Endocr Rev. 2022. PMID: 35596657 Free PMC article. Review.
-
Brain-associated alterations of Hippo pathway transcription in early maturing Atlantic salmon.BMC Ecol Evol. 2025 May 26;25(1):53. doi: 10.1186/s12862-025-02398-4. BMC Ecol Evol. 2025. PMID: 40419966 Free PMC article.
-
The relationship between sperm secretion and the Hippo signaling pathway in the rat seminiferous tubules.Sci Rep. 2025 Aug 2;15(1):28204. doi: 10.1038/s41598-025-12819-5. Sci Rep. 2025. PMID: 40753289 Free PMC article.
References
-
- Bernard DJ, Fortin J, Wang Y, Lamba P. Mechanisms of FSH synthesis: what we know, what we don’t, and why you should care. Fertil Steril. 2010;93(8):2465-2485. - PubMed
-
- Bernard DJ, Tran S. Mechanisms of activin-stimulated FSH synthesis: the story of a pig and a FOX. Biol Reprod. 2013;88(3):78. - PubMed
-
- Kumar TR, Wang Y, Lu N, Matzuk MM. Follicle stimulating hormone is required for ovarian follicle maturation but not male fertility. Nat Genet. 1997;15(2):201-204. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
