

Failures in thymus medulla regeneration during immune recovery cause tolerance loss and prime recipients for auto-GVHD
- PMID: 34910105
- PMCID: PMC8679781
- DOI: 10.1084/jem.20211239
Failures in thymus medulla regeneration during immune recovery cause tolerance loss and prime recipients for auto-GVHD
Abstract
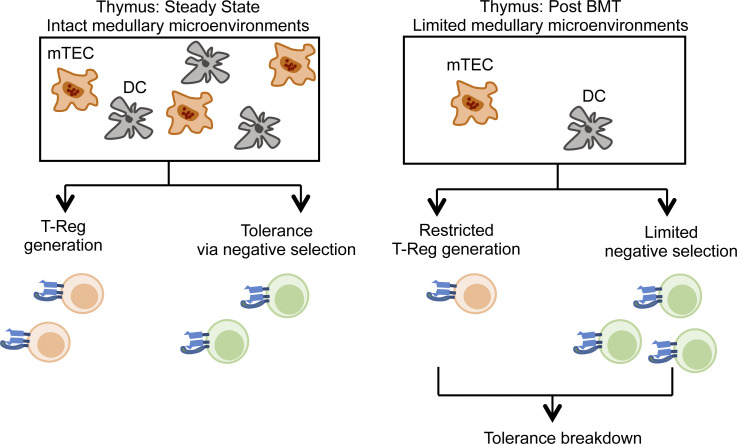
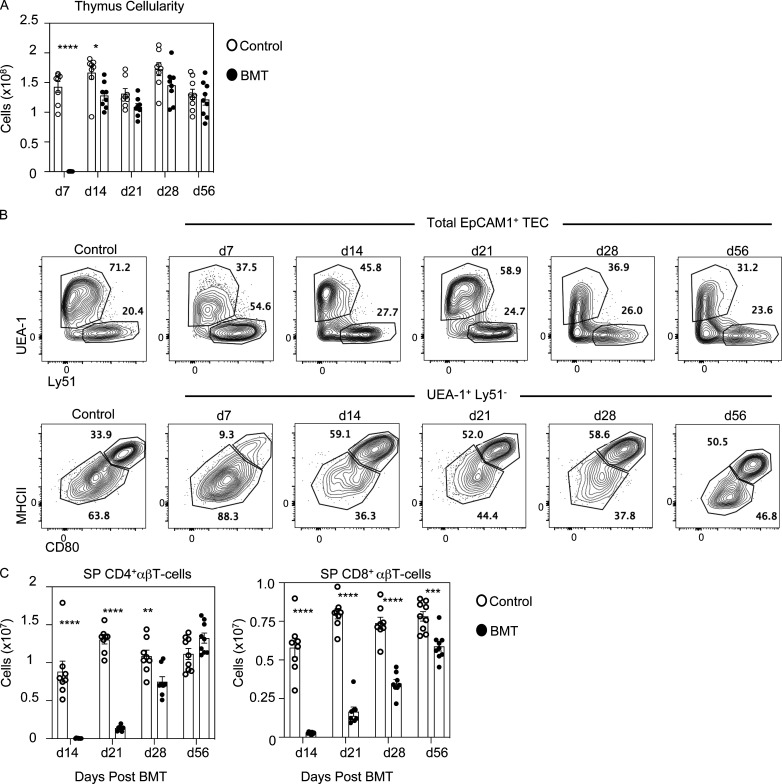
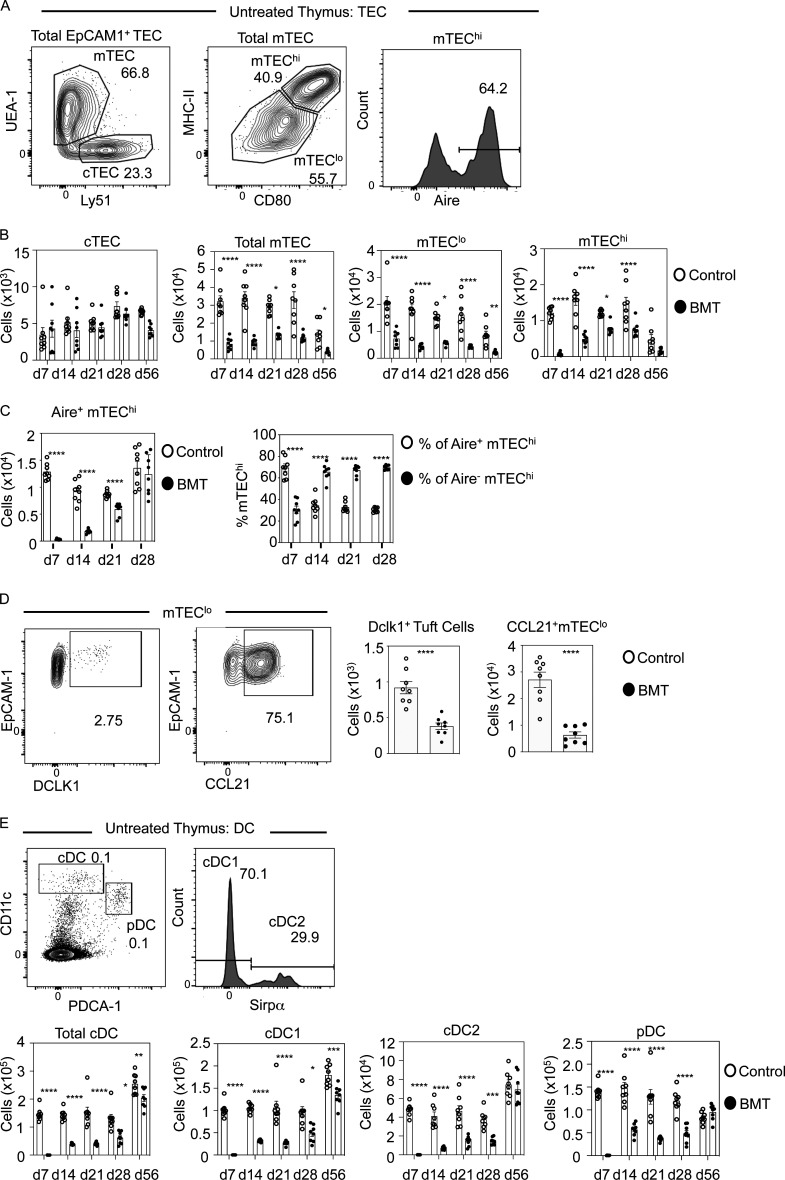
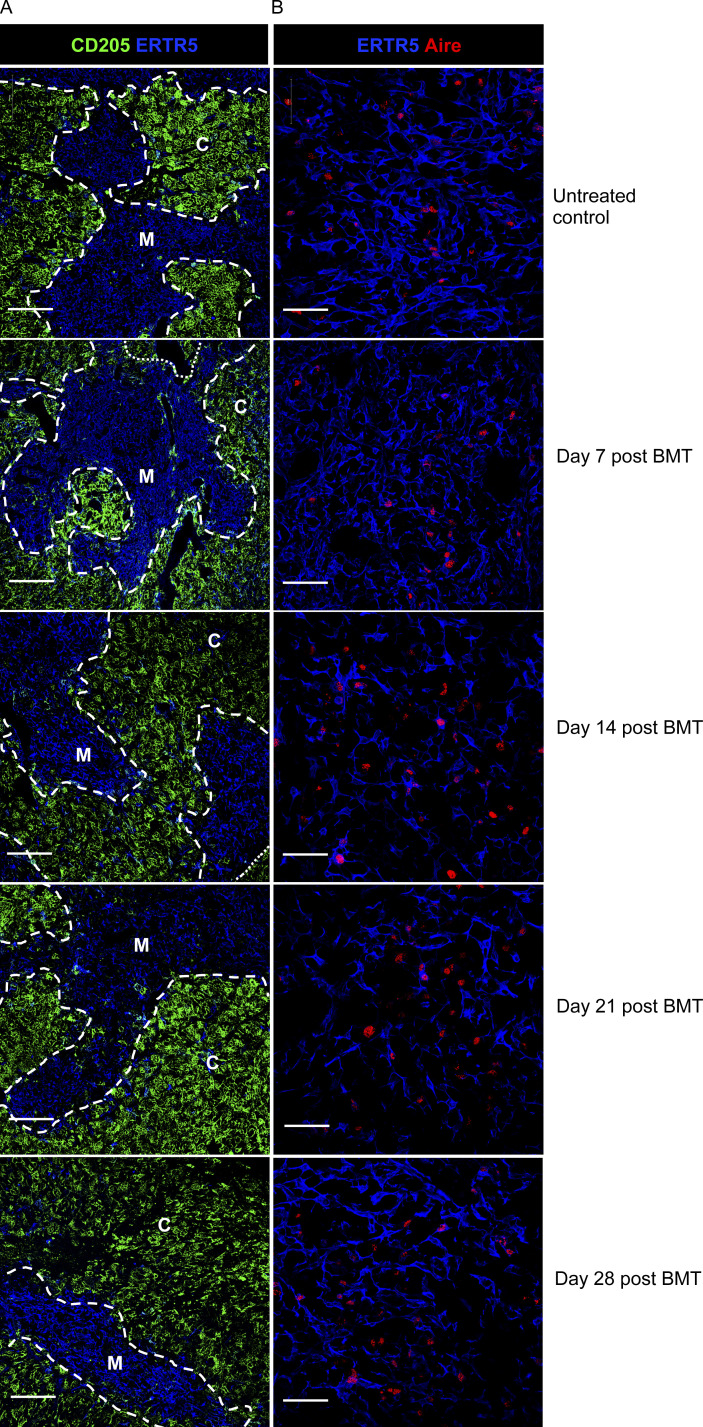
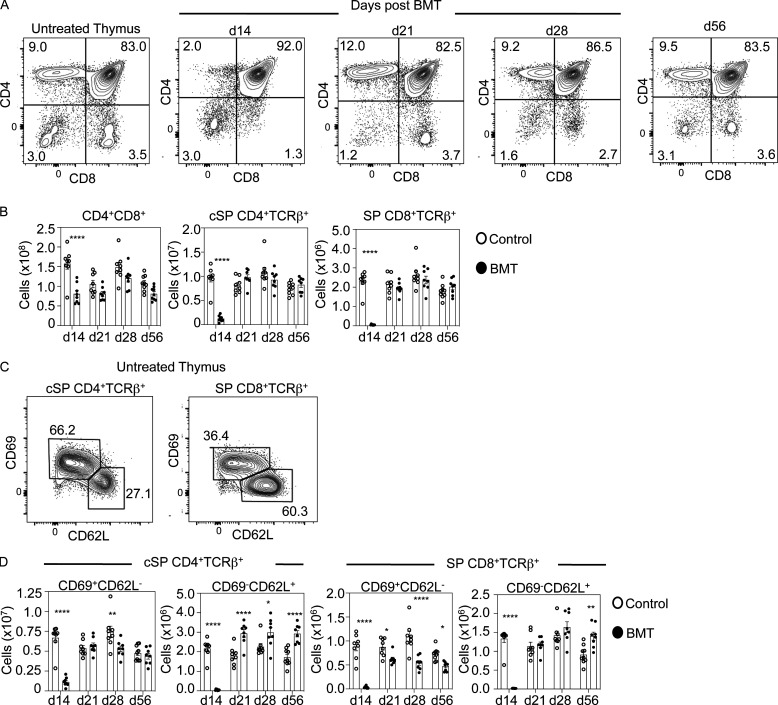
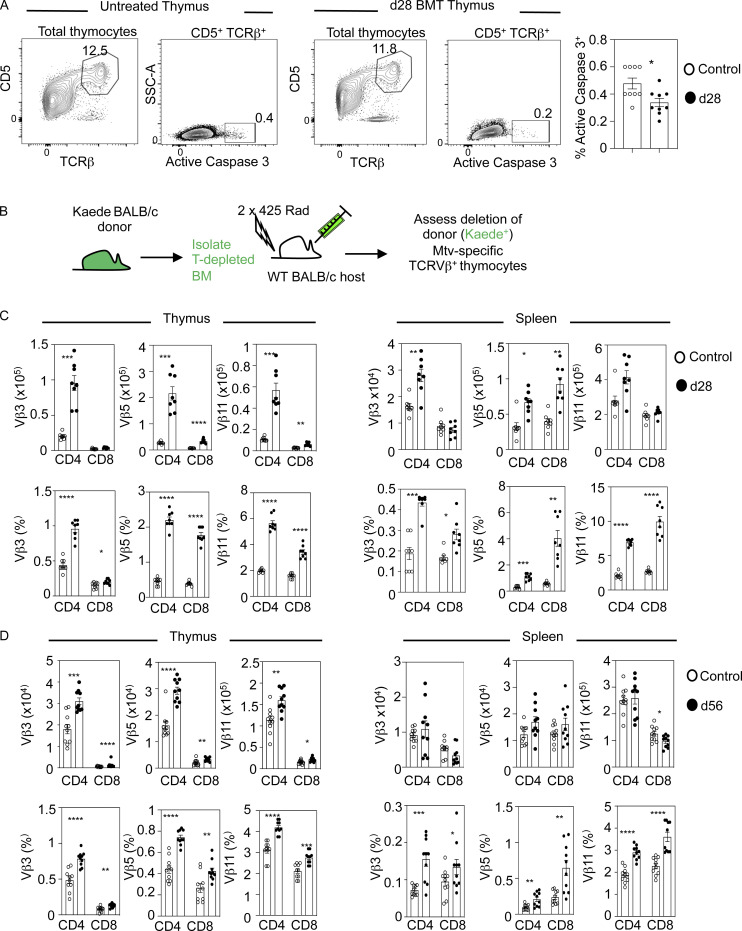
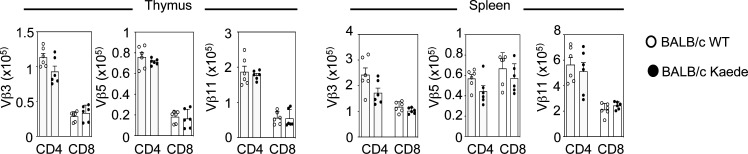
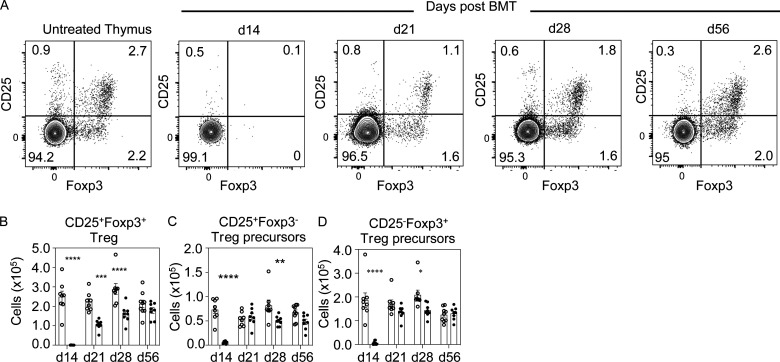
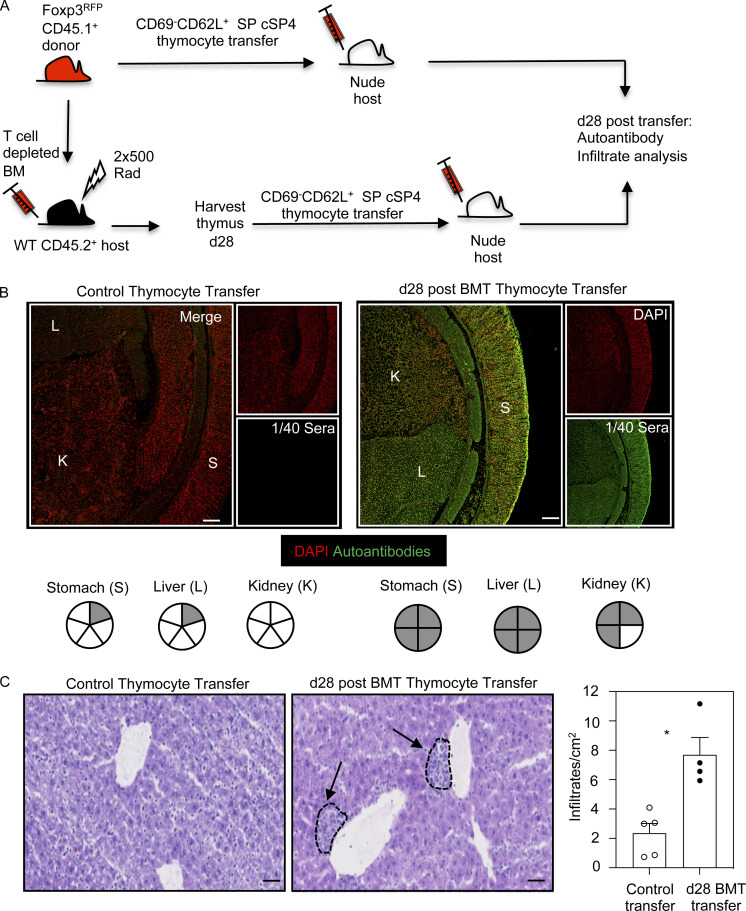
Bone marrow transplantation (BMT) is a widely used therapy for blood cancers and primary immunodeficiency. Following transplant, the thymus plays a key role in immune reconstitution by generating a naive αβT cell pool from transplant-derived progenitors. While donor-derived thymopoiesis during the early post-transplant period is well studied, the ability of the thymus to synchronize T cell development with essential tolerance mechanisms is poorly understood. Using a syngeneic mouse transplant model, we analyzed T cell recovery alongside the regeneration and function of intrathymic microenvironments. We report a specific and prolonged failure in the post-transplant recovery of medullary thymic epithelial cells (mTECs). This manifests as loss of medulla-dependent tolerance mechanisms, including failures in Foxp3+ regulatory T cell development and formation of the intrathymic dendritic cell pool. In addition, defective negative selection enables escape of self-reactive conventional αβT cells that promote autoimmunity. Collectively, we show that post-transplant T cell recovery involves an uncoupling of thymopoiesis from thymic tolerance, which results in autoimmune reconstitution caused by failures in thymic medulla regeneration.
© 2021 Alawam et al.
Conflict of interest statement
Disclosures: The authors declare no competing interests exist.
Figures
Comment in
-
mTEC damage risks immune recovery.J Exp Med. 2022 Feb 7;219(2):e20212326. doi: 10.1084/jem.20212326. Epub 2021 Dec 17. J Exp Med. 2022. PMID: 34919145 Free PMC article.
References
-
- Awong, G., Singh J., Mohtashami M., Malm M., La Motte-Mohs R.N., Benveniste P.M., Serra P., Herer E., van den Brink M.R., and Zúñiga-Pflücker J.C.. 2013. Human proT-cells generated in vitro facilitate hematopoietic stem cell-derived T-lymphopoiesis in vivo and restore thymic architecture. Blood. 122:4210–4219. 10.1182/blood-2012-12-472803 - DOI - PMC - PubMed
-
- Barton-Burke, M., Dwinell D.M., Kafkas L., Lavalley C., Sands H., Proctor C., and Johnson E.. 2008. Graft-versus-host disease: a complex long-term side effect of hematopoietic stem cell transplant. Oncology (Williston Park). 22(11, Suppl Nurse Ed):31–45. - PubMed
