

Localization, Assembly, and Activation of the Escherichia coli Cell Division Machinery
- PMID: 34910577
- PMCID: PMC8919703
- DOI: 10.1128/ecosalplus.ESP-0022-2021
Localization, Assembly, and Activation of the Escherichia coli Cell Division Machinery
Abstract
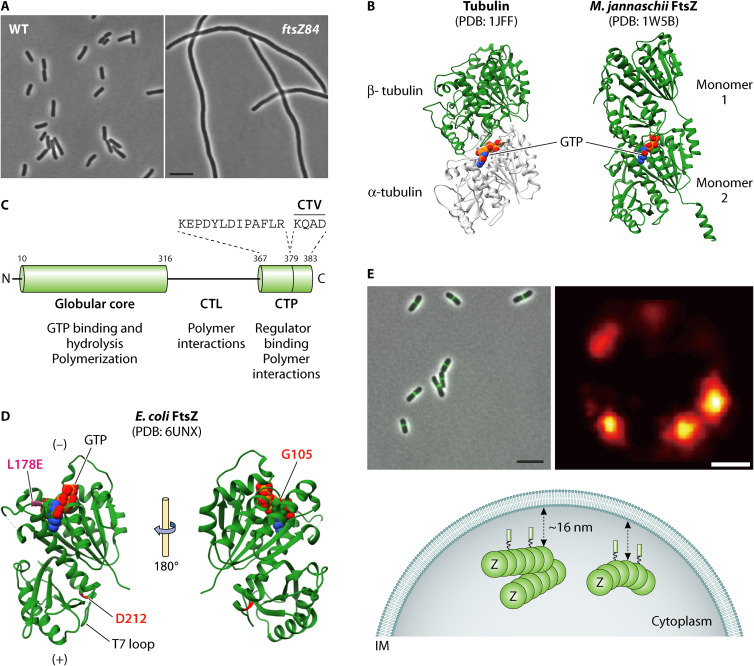
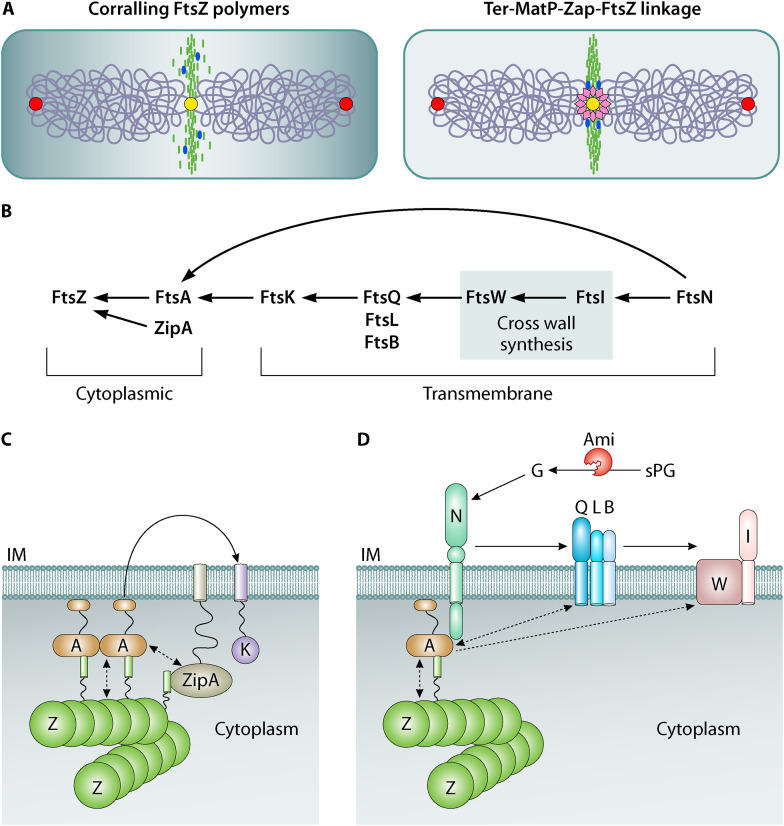
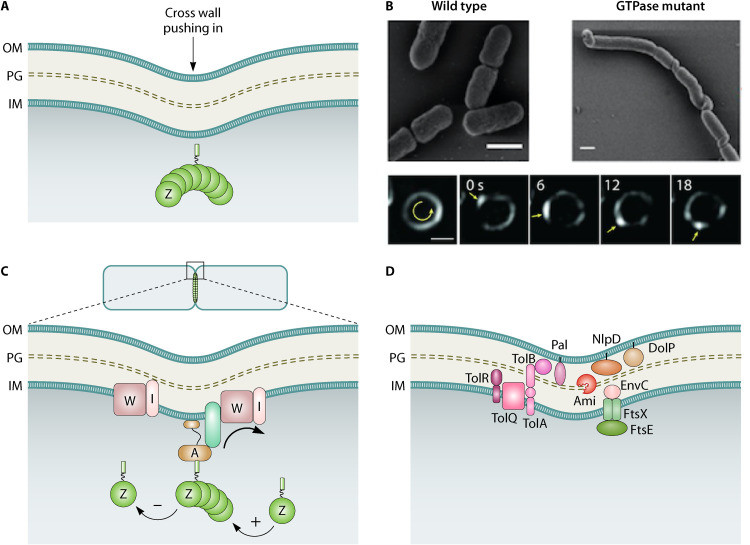
Decades of research, much of it in Escherichia coli, have yielded a wealth of insight into bacterial cell division. Here, we provide an overview of the E. coli division machinery with an emphasis on recent findings. We begin with a short historical perspective into the discovery of FtsZ, the tubulin homolog that is essential for division in bacteria and archaea. We then discuss assembly of the divisome, an FtsZ-dependent multiprotein platform, at the midcell septal site. Not simply a scaffold, the dynamic properties of polymeric FtsZ ensure the efficient and uniform synthesis of septal peptidoglycan. Next, we describe the remodeling of the cell wall, invagination of the cell envelope, and disassembly of the division apparatus culminating in scission of the mother cell into two daughter cells. We conclude this review by highlighting some of the open questions in the cell division field, emphasizing that much remains to be discovered, even in an organism as extensively studied as E. coli.
Keywords: Escherichia coli; FtsZ; cell division; divisome; peptidoglycan.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
